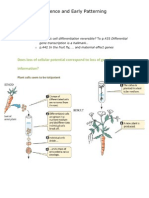
You might also like
- The Pyridine Nucleotide CoenzymesFrom EverandThe Pyridine Nucleotide CoenzymesJohannes EverseNo ratings yet
- Membrane Models: Gorter and Grendel's Membrane Theory (1920)Document5 pagesMembrane Models: Gorter and Grendel's Membrane Theory (1920)Hafiz AhmadNo ratings yet
- Polytene and Lampbrush ChromosomesDocument3 pagesPolytene and Lampbrush Chromosomesneeru.bhagatNo ratings yet
- Molecular PhylogeneticsDocument4 pagesMolecular PhylogeneticsAb WassayNo ratings yet
- Genome Size and Structure, BacterialDocument7 pagesGenome Size and Structure, BacterialCINDY NATHALIA GARCIA AGUDELONo ratings yet
- Genetic Terminology ExplainedDocument6 pagesGenetic Terminology ExplainedMuthu KaruppanNo ratings yet
- BioSci 101 Test 2 NotesDocument12 pagesBioSci 101 Test 2 NotesPianomanSuperman100% (1)
- TransposonesDocument6 pagesTransposonesPiero PaoloNo ratings yet
- PFAM DatabaseDocument22 pagesPFAM DatabaseNadish KumarNo ratings yet
- Angiosperm Classification and The APGDocument2 pagesAngiosperm Classification and The APGAnsev CivasNo ratings yet
- Pieris Homo Lathyrus Sapiens OdoratusDocument1 pagePieris Homo Lathyrus Sapiens OdoratusjennieNo ratings yet
- Lec# Crop Improvement MethodsDocument24 pagesLec# Crop Improvement MethodsSohail AhmedNo ratings yet
- Comparing DNA Sequences To Understand Evolutionary Relationships With BlastDocument3 pagesComparing DNA Sequences To Understand Evolutionary Relationships With Blastflyawayxx13No ratings yet
- Molecular Systematic of AnimalsDocument37 pagesMolecular Systematic of AnimalsMarlene Tubieros - InducilNo ratings yet
- Sequence Alignment: InterpretationDocument8 pagesSequence Alignment: InterpretationdahiyatejNo ratings yet
- Crop ImprovementDocument14 pagesCrop ImprovementhappydaysvsNo ratings yet
- Microbial Genetics & MutationDocument49 pagesMicrobial Genetics & MutationRenz Francis SasaNo ratings yet
- Unit 1: Structural GenomicsDocument4 pagesUnit 1: Structural GenomicsLavanya ReddyNo ratings yet
- Modes of Nutrition in Bacteria ?Document7 pagesModes of Nutrition in Bacteria ?Atika ZulfiqarNo ratings yet
- Transposons (Jumping Genes)Document4 pagesTransposons (Jumping Genes)Aditya Kanwal100% (1)
- Van - Valen - Homology and CausesDocument8 pagesVan - Valen - Homology and CausesJulio Torres100% (1)
- Species InteractionDocument19 pagesSpecies InteractionStyrich Nyl AbayonNo ratings yet
- Exploring Vertebrate EvolutionDocument12 pagesExploring Vertebrate EvolutionPamee BautistaNo ratings yet
- Bioprocess Considerations in Using Animal Cell CulturesDocument25 pagesBioprocess Considerations in Using Animal Cell CulturesDidem Kara100% (1)
- M - 23 Polytene ChromosomeDocument3 pagesM - 23 Polytene ChromosomeDr. Tapan Kr. DuttaNo ratings yet
- Abstracts For 32nd Congrass of ZoologyDocument350 pagesAbstracts For 32nd Congrass of ZoologySaima WaseemNo ratings yet
- 4 Gene ExpressionDocument35 pages4 Gene ExpressionThảo ThảoNo ratings yet
- Genome Organization in ProkaryotesDocument8 pagesGenome Organization in ProkaryotesVijay Kishore75% (4)
- Mitosis and MeiosisDocument13 pagesMitosis and Meiosis420192420192No ratings yet
- Bacterial TransposonsDocument36 pagesBacterial TransposonsShweta SinghNo ratings yet
- Karyotype Analysis in Zea Mays L. Var. Everta (Popcorn) Cultivated Within Owerri, Southeast NigeriaDocument6 pagesKaryotype Analysis in Zea Mays L. Var. Everta (Popcorn) Cultivated Within Owerri, Southeast NigeriaImpact JournalsNo ratings yet
- Bernays & Chapman 1994Document325 pagesBernays & Chapman 1994Alexandre Pimenta100% (2)
- IB Biology HL/SL Auxins and Plant Growth ActivityDocument2 pagesIB Biology HL/SL Auxins and Plant Growth ActivityINDRANI GOSWAMINo ratings yet
- Approval Sheet: Name: Andi Nurlaeli Reg - No: 071404159 Group: IV (Fourth)Document12 pagesApproval Sheet: Name: Andi Nurlaeli Reg - No: 071404159 Group: IV (Fourth)Lhely Andi NurlaeliNo ratings yet
- Assignment 8 Biology EssayDocument6 pagesAssignment 8 Biology Essayjanna conawayNo ratings yet
- Genetics of flower development in Arabidopsis thalianaDocument40 pagesGenetics of flower development in Arabidopsis thalianaSanthoshNo ratings yet
- Cytogenetic Slide PreparationDocument9 pagesCytogenetic Slide PreparationRicardo PalmaNo ratings yet
- Gregor MendelDocument3 pagesGregor MendelAimee GutierrezNo ratings yet
- Eukaryotic Genome Complexity - Learn Science at ScitableDocument4 pagesEukaryotic Genome Complexity - Learn Science at ScitableAnder ManaresNo ratings yet
- Lab 1 Cell and OsmosisDocument5 pagesLab 1 Cell and OsmosisyuwarajaNo ratings yet
- Population Formation by HybridisationDocument50 pagesPopulation Formation by Hybridisationvarsha pNo ratings yet
- Signal Transduction in Cells: An Overview of Key ConceptsDocument25 pagesSignal Transduction in Cells: An Overview of Key ConceptsSadaf BegNo ratings yet
- Genomic Equivalence and Early PatterningDocument11 pagesGenomic Equivalence and Early PatterningSavannah Simone PetrachenkoNo ratings yet
- Silva, Joseph Paulo L. BIO 101 C-2 03/05/2017: Central Dogma of Molecular BiologyDocument3 pagesSilva, Joseph Paulo L. BIO 101 C-2 03/05/2017: Central Dogma of Molecular BiologyJoseph Paulo L SilvaNo ratings yet
- PCR Based Molecualr, Genetic MarkersDocument59 pagesPCR Based Molecualr, Genetic MarkersRafay MirzaNo ratings yet
- Transcription FactorsDocument25 pagesTranscription FactorsPriya.RNo ratings yet
- D. melanogaster Productivity Increased with Banana and Pear EnrichmentDocument6 pagesD. melanogaster Productivity Increased with Banana and Pear EnrichmentEdgar A. ÇolmenárezNo ratings yet
- Phylogeny AnalysisDocument49 pagesPhylogeny AnalysisQuyen Nguyen Thi AnhNo ratings yet
- Anterior Posterior Axis Patterning in DrosophilaDocument4 pagesAnterior Posterior Axis Patterning in DrosophilaSuresh Babu TNo ratings yet
- Genetics and Variation-1Document53 pagesGenetics and Variation-1Hashim SelashiNo ratings yet
- Ascomycota Major Characteristics and LineagesDocument9 pagesAscomycota Major Characteristics and Lineagestiara100% (1)
- Transposable Elements in EukaryotesDocument13 pagesTransposable Elements in EukaryotesAkhilesh BhuraNo ratings yet
- 7 Linkage MappingDocument86 pages7 Linkage MappingEmi JiHyeon KimNo ratings yet
- Gene Regulation in Prokaryotes-1Document17 pagesGene Regulation in Prokaryotes-1Fasiha Mushadi100% (1)
- Movement and Muscle MechanismsDocument12 pagesMovement and Muscle Mechanismsadithya4rajNo ratings yet
- Eukaryotic Protein Synthesis Differs From Prokaryotic Protein Synthesis Primarily in Translation InitiationDocument4 pagesEukaryotic Protein Synthesis Differs From Prokaryotic Protein Synthesis Primarily in Translation InitiationJoyce DometitaNo ratings yet
- Evolution Test CH 10 and 11 Review Sheet ANSWERSDocument4 pagesEvolution Test CH 10 and 11 Review Sheet ANSWERSAlex IoannouNo ratings yet
- FilogeografiaDocument21 pagesFilogeografiaRenata PerezNo ratings yet
- Evans JD Wheeler DE Gene Expression and The Evolution of Insect Polyphenisms BioEssays 23 62 68Document7 pagesEvans JD Wheeler DE Gene Expression and The Evolution of Insect Polyphenisms BioEssays 23 62 68Melivea Paez HerediaNo ratings yet
- Historical Section of PlacentaDocument5 pagesHistorical Section of PlacentaManisha BishtNo ratings yet
- Developmental Biology: Fundamental Processes of DevelopmentDocument9 pagesDevelopmental Biology: Fundamental Processes of DevelopmentManisha BishtNo ratings yet
- 2c. Egg MembranesDocument11 pages2c. Egg MembranesManisha BishtNo ratings yet
- 2b. Types of Egg and CleavageDocument55 pages2b. Types of Egg and CleavageManisha BishtNo ratings yet
- 2b. Types of Egg and CleavageDocument55 pages2b. Types of Egg and CleavageManisha BishtNo ratings yet
- Natural Selection PDFDocument39 pagesNatural Selection PDFManisha BishtNo ratings yet
- Evolution:: The Origin and Evolution of ManDocument39 pagesEvolution:: The Origin and Evolution of ManManisha Bisht100% (4)
- Evidences of Evolution: Geography, Anatomy and FossilsDocument69 pagesEvidences of Evolution: Geography, Anatomy and FossilsManisha Bisht100% (1)
- Orthologs, Paralogs and Evolutionary GenomicsDocument32 pagesOrthologs, Paralogs and Evolutionary GenomicsVIDHI JAINENDRASINH RANANo ratings yet
- Production of Recombinant Pharmaceutical ProteinsDocument25 pagesProduction of Recombinant Pharmaceutical ProteinsJuliana DiazNo ratings yet
- F4 Chapter 5 KSSM SCIENCE DLP EditedDocument138 pagesF4 Chapter 5 KSSM SCIENCE DLP Editedsalizamatisa100% (1)
- Yeast Agglutinin Interaction Characterized at Molecular LevelDocument8 pagesYeast Agglutinin Interaction Characterized at Molecular Levelhaddig8No ratings yet
- U Tube ExperimentDocument143 pagesU Tube ExperimentAbhishek SinghNo ratings yet
- Population Genetics Notes PDFDocument357 pagesPopulation Genetics Notes PDFKaran AshishNo ratings yet
- Quizlet Chapter 1Document6 pagesQuizlet Chapter 1EUNAH LimNo ratings yet
- 1.3 Cell Division and MitosisDocument35 pages1.3 Cell Division and MitosisJay XieNo ratings yet
- Issa Dna Based Fitness Coach Chapter PreviewDocument19 pagesIssa Dna Based Fitness Coach Chapter PreviewUsama Jutt100% (2)
- Psych 372 S'22 Study Guide For QUIZZES 1-3 & EXAM 1Document4 pagesPsych 372 S'22 Study Guide For QUIZZES 1-3 & EXAM 1darby hepplerNo ratings yet
- SRI VENKATESWARA UNIVERSITY MSc Biochemistry SyllabusDocument29 pagesSRI VENKATESWARA UNIVERSITY MSc Biochemistry SyllabusQwertyuNo ratings yet
- What It Means to Be 98% ChimpanzeeDocument328 pagesWhat It Means to Be 98% ChimpanzeeJGokz100% (3)
- Molecular Basis of InheritanceDocument68 pagesMolecular Basis of Inheritancealihaidar23001No ratings yet
- Genetic Engineering EssayDocument4 pagesGenetic Engineering EssayGrantSkroveNo ratings yet
- TaqMan - Life TechnologiesDocument20 pagesTaqMan - Life TechnologiesAni IoanaNo ratings yet
- Cell Growth, Division, and Reproduction Class DateDocument11 pagesCell Growth, Division, and Reproduction Class DateNadim HammoudNo ratings yet
- Biotechnology - Principles and Processes - CompressedDocument113 pagesBiotechnology - Principles and Processes - CompressedPOT SONGNo ratings yet
- Darin Surka 2008Document89 pagesDarin Surka 2008elisaNo ratings yet
- WMSTU Bio2 Course Genetics Cell Cycle Mitosis MeiosisDocument5 pagesWMSTU Bio2 Course Genetics Cell Cycle Mitosis MeiosisCobe Christian LascunaNo ratings yet
- Cell DivisionDocument4 pagesCell DivisionMhariane MabborangNo ratings yet
- Assignment 3 ArkhangelskiiDocument4 pagesAssignment 3 ArkhangelskiiEvgeniiNo ratings yet
- Basic Principles and Components of PCRDocument30 pagesBasic Principles and Components of PCRDiahNo ratings yet
- Economic Traits of PoultryDocument17 pagesEconomic Traits of PoultrySantosh Dhakal100% (2)
- Miranda, Elizabeth S. - BSHM3B (GEC107 W6&W7)Document6 pagesMiranda, Elizabeth S. - BSHM3B (GEC107 W6&W7)Grace Hermano ManiponNo ratings yet
- DNA Evidence Collection GuidelinesDocument4 pagesDNA Evidence Collection GuidelinesJames Van Arquero NaronNo ratings yet
- Reduction of Global H3K27me Enhances HER2ErbB2 Targeted Therapy2019Cell ReportsOpen Access PDFDocument18 pagesReduction of Global H3K27me Enhances HER2ErbB2 Targeted Therapy2019Cell ReportsOpen Access PDFLorena RamosNo ratings yet
- What Are Introns and Exons - PDFDocument14 pagesWhat Are Introns and Exons - PDFSterlingNo ratings yet
- Cte STD Principles Food ProductionDocument6 pagesCte STD Principles Food Productionryvnnnjovu13No ratings yet
- Diferensiasi Dan Spesialisasi SelDocument41 pagesDiferensiasi Dan Spesialisasi SelhusnilwardiyahNo ratings yet
- Lab-2 Genet Ic DNA Isolation Methods-1Document27 pagesLab-2 Genet Ic DNA Isolation Methods-1Samyak ShahNo ratings yet