You might also like
- Hopkins, Tom - How To Master The Art of Selling NEW PDFDocument53 pagesHopkins, Tom - How To Master The Art of Selling NEW PDFDaneMcFarlane67% (6)
- Basant Agrawal C M Agrawal Engineering DrawingDocument778 pagesBasant Agrawal C M Agrawal Engineering Drawingjay singhvi100% (9)
- Basant Agrawal C M Agrawal Engineering DrawingDocument778 pagesBasant Agrawal C M Agrawal Engineering Drawingjay singhvi100% (9)
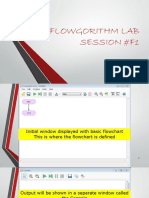
- Flowgorithm PDFDocument56 pagesFlowgorithm PDFKothapalli Likhith Sri SaiNo ratings yet
- Flowgorithm PDFDocument56 pagesFlowgorithm PDFKothapalli Likhith Sri SaiNo ratings yet
- Basant Agrawal C M Agrawal Engineering DrawingDocument778 pagesBasant Agrawal C M Agrawal Engineering Drawingjay singhvi100% (9)
- Sensacion PerceptualDocument219 pagesSensacion PerceptualcircozombieNo ratings yet
- A Nervous TripDocument12 pagesA Nervous TripMark Owen BaldoNo ratings yet
- Shivering Thermogenesis in Humans Origin, Contribution and MetabolicDocument10 pagesShivering Thermogenesis in Humans Origin, Contribution and MetabolicGio VandaNo ratings yet
- Regulation of Body Temperature by The Nervous System PDFDocument18 pagesRegulation of Body Temperature by The Nervous System PDFEstrella BenitoNo ratings yet
- Trashed-0000318Document5 pagesTrashed-0000318Manish GhimireNo ratings yet
- Central Mechanisms of ThermoregulationDocument9 pagesCentral Mechanisms of ThermoregulationYonita RahmiNo ratings yet
- Heldmaier Et Al 1989 Photoperiod and Thermoregulation in Vertebrates Body Temperature Rhythms and ThermogenicDocument15 pagesHeldmaier Et Al 1989 Photoperiod and Thermoregulation in Vertebrates Body Temperature Rhythms and Thermogenicsaryasingh253No ratings yet
- Role of the Preoptic-Anterior Hypothalamus in Thermoregulation and Fever RegulationDocument5 pagesRole of the Preoptic-Anterior Hypothalamus in Thermoregulation and Fever RegulationlutfianaNo ratings yet
- How Neurophysiology Accounts for Thermal Sensation Despite EntropyDocument4 pagesHow Neurophysiology Accounts for Thermal Sensation Despite Entropyfernando_mart859243No ratings yet
- Fever, Temperature, and The Immune Response: DepartmentDocument12 pagesFever, Temperature, and The Immune Response: DepartmentClara OhNo ratings yet
- Perioperative Shivering ReviewDocument18 pagesPerioperative Shivering Reviewwanoe02No ratings yet
- Body Temperature Influences LifespanDocument15 pagesBody Temperature Influences LifespanNomalingeNo ratings yet
- How Plants Sense and Respond To Stressful Environments (Plant Physiology Article)Document12 pagesHow Plants Sense and Respond To Stressful Environments (Plant Physiology Article)Noel CambayaNo ratings yet
- Neural Substrate of Cold-Seeking Behavior in Endotoxin ShockDocument11 pagesNeural Substrate of Cold-Seeking Behavior in Endotoxin ShockGurumoorthy KrishnamoorthyNo ratings yet
- Update Abiotic StressDocument12 pagesUpdate Abiotic StressYuánArgaNo ratings yet
- Coding Efficiency of Fly Motion Processing Is Set by Firing Rate, Not Firing PrecisionDocument11 pagesCoding Efficiency of Fly Motion Processing Is Set by Firing Rate, Not Firing PrecisionspavieriNo ratings yet
- Molecular and Cellular Endocrinology-Thermoregulation in FishDocument8 pagesMolecular and Cellular Endocrinology-Thermoregulation in FishHAROLD AQUILINO RUIZ MOZOMBITENo ratings yet
- Congeneric Variability in Lifespan Extension and Onset of Senescence Suggest Active Regulation of Aging in Response To Low TemperatureDocument8 pagesCongeneric Variability in Lifespan Extension and Onset of Senescence Suggest Active Regulation of Aging in Response To Low Temperatureender000No ratings yet
- Hipotálamo - Control de La Temperatura - Morrison 2011Document47 pagesHipotálamo - Control de La Temperatura - Morrison 2011fede AssirioNo ratings yet
- Physiological Mechanisms of Thermoregulation in ReptilesDocument9 pagesPhysiological Mechanisms of Thermoregulation in ReptilesSalvador ReyesNo ratings yet
- Contribution of Thermal and Nonthermal Factors To The Regulation of BodyDocument8 pagesContribution of Thermal and Nonthermal Factors To The Regulation of BodyMike WazowskiNo ratings yet
- Homeostasis and Feedback Mechanism PDFDocument3 pagesHomeostasis and Feedback Mechanism PDFjer montillaNo ratings yet
- Thermoregulation in Vertebrates: Acclimation, Acclimatization and AdaptationDocument8 pagesThermoregulation in Vertebrates: Acclimation, Acclimatization and Adaptationtrunghieu85No ratings yet
- Ajpcell 00040 2004Document11 pagesAjpcell 00040 2004Anibal SalcedoNo ratings yet
- Beyond The Classic Thermoneutral ZoneDocument9 pagesBeyond The Classic Thermoneutral ZonegeoNo ratings yet
- Big 3Document13 pagesBig 3Fitria Ayu LestariNo ratings yet
- Homeothermic Animal Temperature RegulationDocument3 pagesHomeothermic Animal Temperature RegulationMonesa Christy VillanuevaNo ratings yet
- Animals 11 01733 With CoverDocument28 pagesAnimals 11 01733 With CoverNatalia AguasNo ratings yet
- Homeostatic Disturbance of Thermoregulatory Functions in Rats With Chronic FatigueDocument6 pagesHomeostatic Disturbance of Thermoregulatory Functions in Rats With Chronic FatigueG ElegansNo ratings yet
- estrescronicoDepreZORRILLA ZIEHERDocument10 pagesestrescronicoDepreZORRILLA ZIEHERmarielaNo ratings yet
- Variation in The Heat Shock Response and Its Implication For Predicting The Effect of Global Climate Change On Species' Biogeographical Distribution Ranges and Metabolic CostsDocument9 pagesVariation in The Heat Shock Response and Its Implication For Predicting The Effect of Global Climate Change On Species' Biogeographical Distribution Ranges and Metabolic CostspachriqueNo ratings yet
- Acta Physiologica - 2016 - Mrowka - ThermoregulationDocument3 pagesActa Physiologica - 2016 - Mrowka - ThermoregulationSam Skofic (atomichoplite)No ratings yet
- Evaluating Thermoregulation in ReptilesDocument10 pagesEvaluating Thermoregulation in ReptilesSantiago SuarezNo ratings yet
- The Thermoregulatory Theory of Yawning: What We Know From Over 5 Years of ResearchDocument13 pagesThe Thermoregulatory Theory of Yawning: What We Know From Over 5 Years of ResearchAndré AmorimNo ratings yet
- Thermoregulation in BatsDocument45 pagesThermoregulation in BatsBaraa AkeelNo ratings yet
- LAPORAN PRAKTIKUM FISIOLOGI HEWAN "SUHU TUBUHDocument12 pagesLAPORAN PRAKTIKUM FISIOLOGI HEWAN "SUHU TUBUHAnnida LegiNo ratings yet
- Suggestion With AnswerDocument4 pagesSuggestion With AnswerBeautynot2touchNo ratings yet
- Chapperon Et Al 2016Document9 pagesChapperon Et Al 2016Fernando LimaNo ratings yet
- Cavic Chi Oli 2011Document12 pagesCavic Chi Oli 2011Etna VeraguasNo ratings yet
- The Molecular and Cellular Basis of Cold Sensation: David D. MckemyDocument10 pagesThe Molecular and Cellular Basis of Cold Sensation: David D. MckemyAlyanah AloveraNo ratings yet
- Human Thermoregulation and Measurement of Body Temperature in Exercise and Clinical SettingsDocument7 pagesHuman Thermoregulation and Measurement of Body Temperature in Exercise and Clinical SettingsGriffin MulofwaNo ratings yet
- Evidence For Increased Thermogenesis in Female C57Document4 pagesEvidence For Increased Thermogenesis in Female C57Erickson OngNo ratings yet
- 1 s2.0 S2214180419301655 MainDocument7 pages1 s2.0 S2214180419301655 MainMukti WidodoNo ratings yet
- A newsimplified thermoregulatorybioheatmodelforevaluatingthermalDocument9 pagesA newsimplified thermoregulatorybioheatmodelforevaluatingthermalthi.rar3078No ratings yet
- Tissue 1Document2 pagesTissue 165rangsiman.nanNo ratings yet
- Blouin Demers2005Document7 pagesBlouin Demers2005Rafael Loaiza RamírezNo ratings yet
- Ikegami+et+al Endocrinology Vol156 p647-659Document13 pagesIkegami+et+al Endocrinology Vol156 p647-659sunNo ratings yet
- Thermal homeostasis in the newborn puppy: A review of behavioral and physiological responses to hypothermiaDocument12 pagesThermal homeostasis in the newborn puppy: A review of behavioral and physiological responses to hypothermiaHandayaniNo ratings yet
- Parabrachial Neuron Types Categorically Encode Thermoregulation Variables During Heat Defense - 2020Document17 pagesParabrachial Neuron Types Categorically Encode Thermoregulation Variables During Heat Defense - 2020熙槃No ratings yet
- Manuscript SubmitDocument23 pagesManuscript Submitapi-295124915No ratings yet
- Chapter 28-29 PDFDocument45 pagesChapter 28-29 PDFMikaylaNo ratings yet
- Rea T Transfer in Biomedical Engineering and Biotechnology: Proceedings of TheDocument13 pagesRea T Transfer in Biomedical Engineering and Biotechnology: Proceedings of TheHussain Amir AliNo ratings yet
- Lecture 11 Temperature and Thermodynamics Part 1 Concepts NotesDocument18 pagesLecture 11 Temperature and Thermodynamics Part 1 Concepts NotesYuuKohNo ratings yet
- TMP 4813Document8 pagesTMP 4813FrontiersNo ratings yet
- Mendoza 2010Document5 pagesMendoza 2010Maite Villalobos PerezNo ratings yet
- TRPM8 Activation by Menthol, Icilin, and Cold Is Differentially Modulated by Intracellular PHDocument6 pagesTRPM8 Activation by Menthol, Icilin, and Cold Is Differentially Modulated by Intracellular PHAlyanah AloveraNo ratings yet
- Topic 1: ThermoregulationDocument32 pagesTopic 1: Thermoregulation9090 ARYAN PRASADNo ratings yet
- Targeted Temperature Management in Brain Injured  Patients PDFDocument23 pagesTargeted Temperature Management in Brain Injured  Patients PDFCamilo Benavides BurbanoNo ratings yet
- Physiology of ThermoregulationDocument18 pagesPhysiology of Thermoregulationsanti sidabalokNo ratings yet
- Fisiologi Suhu Tubuh Dan Labirin Sebagai Reseptor Keseimbangan Kel.6Document16 pagesFisiologi Suhu Tubuh Dan Labirin Sebagai Reseptor Keseimbangan Kel.6tiaraarisendaNo ratings yet
- Selected Topics in Environmental Biology: Based on the Sessions on Environmental Biology Held at the XXVI International Congress of Physiological Sciences, New Delhi, October 20-26, 1974From EverandSelected Topics in Environmental Biology: Based on the Sessions on Environmental Biology Held at the XXVI International Congress of Physiological Sciences, New Delhi, October 20-26, 1974Balraj BhatiaNo ratings yet
- 4 - Elementary - Signals Slides - UDocument17 pages4 - Elementary - Signals Slides - UKothapalli Likhith Sri SaiNo ratings yet
- 19ECE282 Digital Electronics and Systems Laboratory/19CCE181 Digital Design Laboratory 2020-24 Batch ECE/CCE/CSE Course PlanDocument1 page19ECE282 Digital Electronics and Systems Laboratory/19CCE181 Digital Design Laboratory 2020-24 Batch ECE/CCE/CSE Course PlanKothapalli Likhith Sri SaiNo ratings yet
- Subspaces of Vector Spaces Math 130 Linear AlgebraDocument2 pagesSubspaces of Vector Spaces Math 130 Linear AlgebraKothapalli Likhith Sri SaiNo ratings yet
- DD Lab ManualDocument12 pagesDD Lab ManualKothapalli Likhith Sri SaiNo ratings yet
- Deeksharambh - Batch of 2020 Student Induction Program Dept of Electronics and Communication EngineeringDocument1 pageDeeksharambh - Batch of 2020 Student Induction Program Dept of Electronics and Communication EngineeringKothapalli Likhith Sri SaiNo ratings yet
- Sundaram ECE301 Notes PDFDocument115 pagesSundaram ECE301 Notes PDFGulrez MNo ratings yet
- Practice Problems - Metal Drude Model and SemiconductorDocument20 pagesPractice Problems - Metal Drude Model and SemiconductorKothapalli Likhith Sri SaiNo ratings yet
- MSN 551 Notes: Ion Implantation and DiffusionDocument64 pagesMSN 551 Notes: Ion Implantation and Diffusionilg1No ratings yet
- Adaptation to Cold Affects ThermoreceptorsDocument10 pagesAdaptation to Cold Affects ThermoreceptorsKothapalli Likhith Sri SaiNo ratings yet
- Gate Standard BooksDocument1 pageGate Standard BooksKothapalli Likhith Sri SaiNo ratings yet
- Gate Standard Books For EceDocument1 pageGate Standard Books For EceKothapalli Likhith Sri SaiNo ratings yet
- Ion-Implantation (Very Clear Explanation)Document47 pagesIon-Implantation (Very Clear Explanation)Kothapalli Likhith Sri SaiNo ratings yet
- Adaptation to Cold Affects ThermoreceptorsDocument10 pagesAdaptation to Cold Affects ThermoreceptorsKothapalli Likhith Sri SaiNo ratings yet
- Advan 00002 2010 PDFDocument10 pagesAdvan 00002 2010 PDFKothapalli Likhith Sri SaiNo ratings yet
- Physiology of PainDocument35 pagesPhysiology of PainJauhar Firdaus50% (2)
- Notes of CH 7 Control and Coordination - Class 10th Science Study RankersDocument9 pagesNotes of CH 7 Control and Coordination - Class 10th Science Study Rankerssamy0% (1)
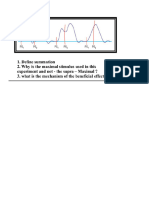
- Summation QTN and AnsDocument5 pagesSummation QTN and AnsaumatonlineNo ratings yet
- FibromyalgiaDocument252 pagesFibromyalgiaAndra Cordun100% (2)
- Role of Perception in Effective CommunicationDocument16 pagesRole of Perception in Effective CommunicationMecha0250% (2)
- General Biology Q4 M7Document19 pagesGeneral Biology Q4 M7Mubin AbdulkarilNo ratings yet
- Science10 Q3 Ver4 Mod3Document31 pagesScience10 Q3 Ver4 Mod3Berliese FriasNo ratings yet
- Classification of Organisms Bio 5090 Notes Chalani WijesuterendraDocument19 pagesClassification of Organisms Bio 5090 Notes Chalani WijesuterendraChal WijeNo ratings yet
- Chapter 1 NotesDocument4 pagesChapter 1 Noteserika sabelNo ratings yet
- Nikola Grahek - Feeling Pain and Being in PainDocument199 pagesNikola Grahek - Feeling Pain and Being in PainАндреа Ковачевић100% (1)
- Chap 02 Set 1 AnswerDocument261 pagesChap 02 Set 1 AnswerKhor Han YiNo ratings yet
- Anatomy and Physiology An Integrative Approach 2nd Edition Mckinley Solutions ManualDocument36 pagesAnatomy and Physiology An Integrative Approach 2nd Edition Mckinley Solutions Manualgavageaiderpnbn87100% (27)
- Marketing ResearchDocument22 pagesMarketing ResearchHyacinth Joy MaldepeñaNo ratings yet
- Maslow's Hierarchy - Use As ReferenceDocument2 pagesMaslow's Hierarchy - Use As ReferenceZarah UdaniNo ratings yet
- FIORITO. Is There Pain in Invertebrates PDFDocument6 pagesFIORITO. Is There Pain in Invertebrates PDFErnesto CastroNo ratings yet
- Individual Aspects of Consumer Perception, Learning & Information ProcessingDocument90 pagesIndividual Aspects of Consumer Perception, Learning & Information ProcessingDeepika AgarwalNo ratings yet
- (1957) An Opponent-Process Theory of Color VisionDocument21 pages(1957) An Opponent-Process Theory of Color Visionjoju0531No ratings yet
- Uts Physical SelfDocument3 pagesUts Physical SelfShane Razal II100% (1)
- THE INTEGRATIVE ACTION OF THE NERVOUS SYSTEM - Charles SherringtonDocument442 pagesTHE INTEGRATIVE ACTION OF THE NERVOUS SYSTEM - Charles SherringtonGastón Tadeo ReyeroNo ratings yet
- Models of Consumer BehaviorDocument7 pagesModels of Consumer BehaviorAnuj BhatnagarNo ratings yet
- CH 3 - Study GuideDocument13 pagesCH 3 - Study Guide2688gieNo ratings yet
- Dopamine, Smartphones & You - A Battle For Your Time - Science in The News PDFDocument18 pagesDopamine, Smartphones & You - A Battle For Your Time - Science in The News PDFLeandroCisternaHernandezNo ratings yet
- Experiment No. 4 Reaction Time For Visual Stimuli With and Without MusicDocument8 pagesExperiment No. 4 Reaction Time For Visual Stimuli With and Without MusicFatima MalikNo ratings yet
- Lesson 3 Nervous SystemDocument11 pagesLesson 3 Nervous SystemMARY JANE ANGELICA SEVANo ratings yet
- A History of Perimetry and Visual Field Testing.5Document8 pagesA History of Perimetry and Visual Field Testing.5petrarizkyNo ratings yet
- Multiple Choice Questions: A. B. C. DDocument37 pagesMultiple Choice Questions: A. B. C. DwanderagroNo ratings yet
- Sensory Deprivation Seminar: Effects and Nursing RoleDocument24 pagesSensory Deprivation Seminar: Effects and Nursing Rolesimonjosan100% (2)