You might also like
- 2023 The - Cortico-Limbo-Thalamo-Cortical - Circuits PDFDocument20 pages2023 The - Cortico-Limbo-Thalamo-Cortical - Circuits PDFUSM San IgnacioNo ratings yet
- Brainstem and Nuclei Condensed Grayscale SlidesDocument17 pagesBrainstem and Nuclei Condensed Grayscale SlidesMobarobberNo ratings yet
- The Pyramidal Tract: New Pathways: Revista de Neurologia June 2001Document9 pagesThe Pyramidal Tract: New Pathways: Revista de Neurologia June 2001daniela andreaNo ratings yet
- DiafragmaDocument8 pagesDiafragmaRodrigo Diaz VqzNo ratings yet
- International Standards For Neurological and Functional Classification of Spinal Cord InjuryDocument14 pagesInternational Standards For Neurological and Functional Classification of Spinal Cord InjuryVijay IyerNo ratings yet
- PNAS2011 Petanjek NEOTsynaptsDocument6 pagesPNAS2011 Petanjek NEOTsynaptsAlejandraNo ratings yet
- Edney 1986Document10 pagesEdney 1986Itai IzhakNo ratings yet
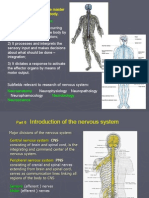
- Three Main Functions:: The Nervous System Is The Master Controlling System of The BodyDocument34 pagesThree Main Functions:: The Nervous System Is The Master Controlling System of The Bodyapi-19641337No ratings yet
- The Anatomical Record - 2013 - Hern Ndez Morato - Somatotopy of The Neurons Innervating The Cricothyroid PosteriorDocument10 pagesThe Anatomical Record - 2013 - Hern Ndez Morato - Somatotopy of The Neurons Innervating The Cricothyroid PosteriormanuelNo ratings yet
- Fernandez-Miranda JC. 2008 - The Claustrum and Its Projection System in The Human BrainDocument11 pagesFernandez-Miranda JC. 2008 - The Claustrum and Its Projection System in The Human BrainBeyza ZeyrekNo ratings yet
- A CanedoDocument15 pagesA CanedoBrenda Espinoza CastilloNo ratings yet
- La Vía Piramidal: Nuevas Trayectorias: M.L. Cuadrado, J.A. Arias, M.A. Palomar, R. LinaresDocument8 pagesLa Vía Piramidal: Nuevas Trayectorias: M.L. Cuadrado, J.A. Arias, M.A. Palomar, R. Linarescl.lexisalazar00No ratings yet
- 001 004 PDFDocument4 pages001 004 PDFPutu Gede SudiraNo ratings yet
- Microsurgical Anatomy of The Central Core of The BrainDocument18 pagesMicrosurgical Anatomy of The Central Core of The BrainZeptalanNo ratings yet
- 1-S2.0-S1878875022005460-Main Filum Terminale: A Comprehensive Review With Anatomical, Pathological, and Surgical ConsiderationDocument10 pages1-S2.0-S1878875022005460-Main Filum Terminale: A Comprehensive Review With Anatomical, Pathological, and Surgical ConsiderationYo MeNo ratings yet
- Trig Emi No VagalDocument11 pagesTrig Emi No Vagaljonathan ahladasNo ratings yet
- 1 PRECUNEUSselfdanceDocument20 pages1 PRECUNEUSselfdancePaola Andrea GomezNo ratings yet
- 666 AOCDraftDocument7 pages666 AOCDraftYusuf BrilliantNo ratings yet
- 1980-Axons From CNS Neurones Regenerate Into PNS GraftsDocument2 pages1980-Axons From CNS Neurones Regenerate Into PNS GraftsWim ZhuNo ratings yet
- The MidbrainDocument3 pagesThe MidbrainAshly Kate AbarientosNo ratings yet
- Rewiew FINAL 1Document13 pagesRewiew FINAL 1achalbob18No ratings yet
- Introduction and Spinal CordDocument27 pagesIntroduction and Spinal Cordapi-196413370% (1)
- Vulliemoz 2005Document13 pagesVulliemoz 2005ShreeNo ratings yet
- The Pyramidal Tract: New Pathways: Revista de Neurologia January 2001Document9 pagesThe Pyramidal Tract: New Pathways: Revista de Neurologia January 2001javier solisNo ratings yet
- General Appearance ThalamusDocument7 pagesGeneral Appearance ThalamusYubi KimNo ratings yet
- Henne Man 1965Document21 pagesHenne Man 1965Brayan BuitragoNo ratings yet
- Hearing Research: Hisayuki Ojima, Kunio MurakamiDocument8 pagesHearing Research: Hisayuki Ojima, Kunio MurakamiFrontiersNo ratings yet
- Neuroanatomy of The Zebrafish Brain - A Topological Atlas 1996Document141 pagesNeuroanatomy of The Zebrafish Brain - A Topological Atlas 1996Daniel RuizNo ratings yet
- What Are The Basal Ganglia?Document53 pagesWhat Are The Basal Ganglia?Gursangeetkaur2010No ratings yet
- Nervous TissueDocument6 pagesNervous TissueAhmed JawdetNo ratings yet
- Cerebellum TextDocument7 pagesCerebellum TextJean Pierre Chastre LuzaNo ratings yet
- Rullida - EdullantesDocument36 pagesRullida - EdullantesJaleah Gwyneth Fernandez EdullantesNo ratings yet
- Human BrainDocument44 pagesHuman BrainSri vaishnava100% (1)
- The Cerebellum Revisited (Marion Wassef, Pierre Angaut, Leonor Arsenio-Nune (B-Ok - Xyz)Document353 pagesThe Cerebellum Revisited (Marion Wassef, Pierre Angaut, Leonor Arsenio-Nune (B-Ok - Xyz)hidrogeolo_29No ratings yet
- Neuropsych Studies of The Frontal LobesDocument26 pagesNeuropsych Studies of The Frontal Lobesстас кшижевскийNo ratings yet
- 2003 Structure and Innervation of The CochleaDocument26 pages2003 Structure and Innervation of The CochleaCaio LeônidasNo ratings yet
- Snell Reticular Formation and Limbic SystemDocument9 pagesSnell Reticular Formation and Limbic SystemFrnz RiveraNo ratings yet
- Introduction and Organization of Nervous System: Presented by Dr. Amen FatimaDocument37 pagesIntroduction and Organization of Nervous System: Presented by Dr. Amen FatimaShanailNo ratings yet
- Hornung 2010Document14 pagesHornung 2010Leandy MartinezNo ratings yet
- Anatomia Plexului BrahialDocument10 pagesAnatomia Plexului BrahialsuzzixxNo ratings yet
- Safari - Feb 21, 2024 at 12:17 PMDocument1 pageSafari - Feb 21, 2024 at 12:17 PMsyansyncNo ratings yet
- Acknowledgments: Gray's Anatomy, The Remarkable Work From Which Gray's Clinical Neuroanatomy IsDocument7 pagesAcknowledgments: Gray's Anatomy, The Remarkable Work From Which Gray's Clinical Neuroanatomy IsAndreea LăzăroiuNo ratings yet
- C&B Chapt 2Document29 pagesC&B Chapt 2apfelbaum11122No ratings yet
- Motor SysDocument72 pagesMotor SysLeoNo ratings yet
- 4.18 The Hominin Fossil Record, Emergence of The Modern CNSDocument46 pages4.18 The Hominin Fossil Record, Emergence of The Modern CNSicarus85No ratings yet
- Lemon 2021 - The Cortical "Upper Motoneuron" in Health and DiseaseDocument10 pagesLemon 2021 - The Cortical "Upper Motoneuron" in Health and DiseaselicmarinamaranNo ratings yet
- Biophysio Prelim ExamsDocument2 pagesBiophysio Prelim ExamsKimNo ratings yet
- Review of Anatomy and Physiology of CNS: Blood-Brain BarrierDocument36 pagesReview of Anatomy and Physiology of CNS: Blood-Brain BarrierTimothy CaragNo ratings yet
- Fernandez-Miranda JC. 2007 - Three-Dimensional Microsurgical and Tractographic Anatomy of The White Matter..Document40 pagesFernandez-Miranda JC. 2007 - Three-Dimensional Microsurgical and Tractographic Anatomy of The White Matter..Beyza ZeyrekNo ratings yet
- The Tail of The Striatum, From Anatomy To Connectivity and Function - Emmanuel Valjent and Giuseppe GangarossaDocument12 pagesThe Tail of The Striatum, From Anatomy To Connectivity and Function - Emmanuel Valjent and Giuseppe GangarossaAku KamuNo ratings yet
- Corticospinal Tract Pyramidal Tract - PhysiopediaDocument4 pagesCorticospinal Tract Pyramidal Tract - PhysiopediaNrs Sani Sule MashiNo ratings yet
- Central Components of The Motorsystemand Clinical Syndromes of Lesions Affecting ThemDocument5 pagesCentral Components of The Motorsystemand Clinical Syndromes of Lesions Affecting ThemItzel RomualdoNo ratings yet
- Oraganisation of The Brain: Studenti (Gr. 13) : - Paun Cristina - Petrescu Andreea Elena - Poenaru Raluca SanzianaDocument10 pagesOraganisation of The Brain: Studenti (Gr. 13) : - Paun Cristina - Petrescu Andreea Elena - Poenaru Raluca SanzianaPaun CristinaNo ratings yet
- Neuroanatomy, Brainstem - StatPearls - NCBI BookshelfDocument13 pagesNeuroanatomy, Brainstem - StatPearls - NCBI BookshelfKritika MalhotraNo ratings yet
- Presentation 3 (Information) Major Descending (Motor) Pathway and Spinal Cord Tracts March 25, 2012Document23 pagesPresentation 3 (Information) Major Descending (Motor) Pathway and Spinal Cord Tracts March 25, 2012bsittNo ratings yet
- Pelkey. HIPPOCAMPAL GABAERGIC INHIBITORY INTERNEURONSDocument129 pagesPelkey. HIPPOCAMPAL GABAERGIC INHIBITORY INTERNEURONSmaariamunoz9No ratings yet
- Neuro AnatomyDocument24 pagesNeuro Anatomybirijik7979No ratings yet
- Ns1 Assignment 1 Neuroembryology and NeurohistologyDocument4 pagesNs1 Assignment 1 Neuroembryology and NeurohistologyMark AbrazaldoNo ratings yet
- CerebellumDocument13 pagesCerebellumNejla HNo ratings yet
- Hugdahl 2005Document17 pagesHugdahl 2005ShreeNo ratings yet
- Whitehead 2013Document17 pagesWhitehead 2013ShreeNo ratings yet
- Laterality: Asymmetries of Body, Brain and CognitionDocument23 pagesLaterality: Asymmetries of Body, Brain and CognitionShreeNo ratings yet
- Mend Lo Vic 2018Document18 pagesMend Lo Vic 2018ShreeNo ratings yet
- Asymmetric Brain Function, Affective Style, and Psychopathology: The Role of Early Experience and PlasticityDocument18 pagesAsymmetric Brain Function, Affective Style, and Psychopathology: The Role of Early Experience and PlasticityShreeNo ratings yet
- Vulliemoz 2005Document13 pagesVulliemoz 2005ShreeNo ratings yet
- Dublin Journal: Medical ScienceDocument4 pagesDublin Journal: Medical ScienceShreeNo ratings yet
- FocusDocument6 pagesFocusZubair Mahmood KamalNo ratings yet
- Neuron Patterns Controlling Transmission of Ipsilateral Hind Limb Reflexesin CATDocument23 pagesNeuron Patterns Controlling Transmission of Ipsilateral Hind Limb Reflexesin CATShreeNo ratings yet
- Multiple Intelligences ChecklistDocument6 pagesMultiple Intelligences ChecklistAnthony ChaiNo ratings yet
- Thematic Apperception TestDocument69 pagesThematic Apperception TestShree100% (1)
- Reaching To Ipsilateral or Contralateral Targets: Within-Hemisphere Visuomotor Processing Cannot Explain Hemispatial Differences in Motor ControlDocument9 pagesReaching To Ipsilateral or Contralateral Targets: Within-Hemisphere Visuomotor Processing Cannot Explain Hemispatial Differences in Motor ControlShreeNo ratings yet
- Measuring Needs With The Thematic Apperception Test: A Psychometric StudyDocument15 pagesMeasuring Needs With The Thematic Apperception Test: A Psychometric StudyShreeNo ratings yet
- Review Article 2Document13 pagesReview Article 2ShreeNo ratings yet
- An Indian Modification of The Thematic Apperception TestDocument20 pagesAn Indian Modification of The Thematic Apperception TestShreeNo ratings yet
- Engler & Brown - Stages of Mindfulness MeditationDocument50 pagesEngler & Brown - Stages of Mindfulness MeditationAndrea TuranoNo ratings yet
- Reena Paper PublishedDocument9 pagesReena Paper PublishedShreeNo ratings yet
- Al-Baraa Ibn Malik Al-AnsariDocument3 pagesAl-Baraa Ibn Malik Al-AnsariRahbarTvNo ratings yet
- AP Online Quiz KEY Chapter 8: Estimating With ConfidenceDocument6 pagesAP Online Quiz KEY Chapter 8: Estimating With ConfidenceSaleha IftikharNo ratings yet
- United States Court of Appeals Fifth CircuitDocument4 pagesUnited States Court of Appeals Fifth CircuitScribd Government DocsNo ratings yet
- Research Article: Old Sagay, Sagay City, Negros Old Sagay, Sagay City, Negros Occidental, PhilippinesDocument31 pagesResearch Article: Old Sagay, Sagay City, Negros Old Sagay, Sagay City, Negros Occidental, PhilippinesLuhenNo ratings yet
- Roger Dean Kiser Butterflies)Document4 pagesRoger Dean Kiser Butterflies)joitangNo ratings yet
- Text Mohamed AliDocument2 pagesText Mohamed AliARYAJAI SINGHNo ratings yet
- Molly On The Shore by Percy Grainger Unit StudyDocument5 pagesMolly On The Shore by Percy Grainger Unit Studyapi-659613441No ratings yet
- Arthropods: Surviving The Frost: Charmayne Roanna L. GalangDocument2 pagesArthropods: Surviving The Frost: Charmayne Roanna L. GalangBabes-Rose GalangNo ratings yet
- FCI - GST - Manual On Returns and PaymentsDocument30 pagesFCI - GST - Manual On Returns and PaymentsAmber ChaturvediNo ratings yet
- Pr1 m4 Identifying The Inquiry and Stating The ProblemDocument61 pagesPr1 m4 Identifying The Inquiry and Stating The ProblemaachecheutautautaNo ratings yet
- Simple FTP UploadDocument10 pagesSimple FTP Uploadagamem1No ratings yet
- Cambridge O Level: Pakistan Studies 2059/02Document4 pagesCambridge O Level: Pakistan Studies 2059/02Azfar RashedNo ratings yet
- UntitledDocument2 pagesUntitledRoger GutierrezNo ratings yet
- PT3 Liste PDFDocument2 pagesPT3 Liste PDFSiti KamalNo ratings yet
- Fish50 Write UpDocument3 pagesFish50 Write UpSophia ManzanoNo ratings yet
- Math 3140-004: Vector Calculus and PdesDocument6 pagesMath 3140-004: Vector Calculus and PdesNathan MonsonNo ratings yet
- Ode To The West Wind Text and AnalysisDocument7 pagesOde To The West Wind Text and AnalysisAbdullah HamzaNo ratings yet
- British Council IELTS Online Application SummaryDocument3 pagesBritish Council IELTS Online Application Summarys_asadeNo ratings yet
- Presbuteroi (Elders) and Episkopoi (Overseers) and Are Described in 1 Tim 3 and TitusDocument15 pagesPresbuteroi (Elders) and Episkopoi (Overseers) and Are Described in 1 Tim 3 and TitusNimaro Brenda100% (1)
- Intro To LodgingDocument63 pagesIntro To LodgingjaevendNo ratings yet
- What's More: Quarter 2 - Module 7: Deferred AnnuityDocument4 pagesWhat's More: Quarter 2 - Module 7: Deferred AnnuityChelsea NicoleNo ratings yet
- SyerynDocument2 pagesSyerynHzlannNo ratings yet
- Disciplinary Literacy Strategies To Support Transactions in Elementary Social StudiesDocument11 pagesDisciplinary Literacy Strategies To Support Transactions in Elementary Social Studiesmissjoseph0803No ratings yet
- Faculty of Computer Science and Information TechnologyDocument4 pagesFaculty of Computer Science and Information TechnologyNurafiqah Sherly Binti ZainiNo ratings yet
- Chapter 1Document8 pagesChapter 1Shidan MohdNo ratings yet
- Chapter 13 Creating and Managing TeamsDocument40 pagesChapter 13 Creating and Managing TeamsH.No ratings yet
- Lozada Vs MendozaDocument4 pagesLozada Vs MendozaHarold EstacioNo ratings yet
- Case Study GingerDocument2 pagesCase Study Gingersohagdas0% (1)
- Solaris Hardening Guide v1Document56 pagesSolaris Hardening Guide v1GusGualdNo ratings yet
- Jao Vs Court of Appeals G.R. No. 128314 May 29, 2002Document3 pagesJao Vs Court of Appeals G.R. No. 128314 May 29, 2002Ma Gabriellen Quijada-TabuñagNo ratings yet