You might also like
- Cold Spring Harb Perspect Biol-2012-Blakely-A005595Document25 pagesCold Spring Harb Perspect Biol-2012-Blakely-A005595Md. Ahsan-Ul BariNo ratings yet
- Vasopressin in Septic ShockDocument8 pagesVasopressin in Septic ShockVaishnavi SheteNo ratings yet
- Synaptic Transmission: Communication Between NeuronsDocument48 pagesSynaptic Transmission: Communication Between NeuronsEarl Adam LambacoNo ratings yet
- Molecular Mechanisms of Neurotransmitter ReleaseDocument21 pagesMolecular Mechanisms of Neurotransmitter ReleaseNusrat JahanNo ratings yet
- Arginine Vasopressin Antagonists For The Treatment of Heart Failure and HyponatremiaDocument12 pagesArginine Vasopressin Antagonists For The Treatment of Heart Failure and Hyponatremiadhimas satriaNo ratings yet
- T3 Biokimia Peptide HormonDocument8 pagesT3 Biokimia Peptide HormonFarhan FadhillahNo ratings yet
- Tra 12692Document16 pagesTra 12692戴翰明No ratings yet
- Regulation of Nephron Progenitor Cell Self-Renewal by Intermediary MetabolismDocument13 pagesRegulation of Nephron Progenitor Cell Self-Renewal by Intermediary MetabolismWilan KrisnaNo ratings yet
- IntestineDocument10 pagesIntestineIoanna PerakiNo ratings yet
- Saqs - (Very Important)Document67 pagesSaqs - (Very Important)BRIGHTON JOSHUANo ratings yet
- MKN 021Document4 pagesMKN 021Andy SwiftNo ratings yet
- Diabetologia: The Cell Biology of The Insulin ReceptorDocument9 pagesDiabetologia: The Cell Biology of The Insulin ReceptorAnwar Hosen OntorNo ratings yet
- Red wine compound relaxes arteries via transport proteinDocument5 pagesRed wine compound relaxes arteries via transport proteinFrancisco RamirezNo ratings yet
- Basic Science Review: Functional Hyperemia in The BrainDocument7 pagesBasic Science Review: Functional Hyperemia in The BraindediNo ratings yet
- Cancer NK Cell ActivationDocument7 pagesCancer NK Cell ActivationMazlan SijifNo ratings yet
- Swelling-Activated CL Channels Support CL Secretion by Bovine Ciliary EpitheliumDocument7 pagesSwelling-Activated CL Channels Support CL Secretion by Bovine Ciliary EpitheliumShashi kant ChaudharyNo ratings yet
- How Neurons Communicate To Each Other at A Chemical Synapse?Document5 pagesHow Neurons Communicate To Each Other at A Chemical Synapse?stepkim92No ratings yet
- Avances en Neurobiologia de La Union Neuromusuclar Anestesiology 2002Document30 pagesAvances en Neurobiologia de La Union Neuromusuclar Anestesiology 2002Irene ChimalNo ratings yet
- Lovely Professional University: Assignment ReportDocument5 pagesLovely Professional University: Assignment ReportAdarshNo ratings yet
- Mikula Phosphorylation2006Document8 pagesMikula Phosphorylation2006julia rossoNo ratings yet
- Nrneph 2010 157Document10 pagesNrneph 2010 157spraptamaNo ratings yet
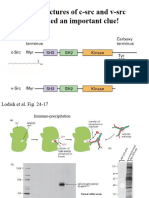
- The Structures of C-SRC and V-SRC Provided An Important Clue!Document33 pagesThe Structures of C-SRC and V-SRC Provided An Important Clue!Souvik DeNo ratings yet
- ARTICULO 1cell Signallingcornell2005Document4 pagesARTICULO 1cell Signallingcornell2005Carmen MaRiNo ratings yet
- Nav1.7 Sodium Channel Isoform Is Expressed in HippocampalDocument7 pagesNav1.7 Sodium Channel Isoform Is Expressed in HippocampalGiriraj SahuNo ratings yet
- Contribution of Retinoid X Receptor Signaling To The Specification of Skeletal Muscle LineageDocument7 pagesContribution of Retinoid X Receptor Signaling To The Specification of Skeletal Muscle LineageFrontiersNo ratings yet
- (Hua et al. 2010) A mechanosensitive ion channel regulating cell volumeDocument7 pages(Hua et al. 2010) A mechanosensitive ion channel regulating cell volumektmahbubaniNo ratings yet
- Cell CycleDocument2 pagesCell CycleИгорьNo ratings yet
- The Role of Pannexin-1 Channel - ReviewDocument4 pagesThe Role of Pannexin-1 Channel - ReviewInternational Journal of Innovative Science and Research TechnologyNo ratings yet
- CXCL12 PDFDocument17 pagesCXCL12 PDFJean andre CanoNo ratings yet
- Apoptosis 4Document16 pagesApoptosis 4Kamila Środa-PomianekNo ratings yet
- Specific Binding of RBC To EC Is Regulated by Nonadsorbing MacromoleculesDocument7 pagesSpecific Binding of RBC To EC Is Regulated by Nonadsorbing MacromoleculesaguslistyNo ratings yet
- Activation of A Nadph Oxidase From Horse Polymorphonuclear Leukocytes in A Cell-Free SystemDocument9 pagesActivation of A Nadph Oxidase From Horse Polymorphonuclear Leukocytes in A Cell-Free SystemDuaa ShamiehNo ratings yet
- 12 Boute 00 NatGenetDocument6 pages12 Boute 00 NatGenetDorin Cristian AntalNo ratings yet
- Biological Relevance of Neurotransmitter ReleaseDocument12 pagesBiological Relevance of Neurotransmitter ReleaseazzaassNo ratings yet
- Central Clock Excites Vasopressin Neurons by Waking Osmosensory Afferents During Late SleepDocument9 pagesCentral Clock Excites Vasopressin Neurons by Waking Osmosensory Afferents During Late SleepLydia BoumansourNo ratings yet
- Effects of Collagen IV On Neuroblastoma Cell Matrix-Related FunctionsDocument9 pagesEffects of Collagen IV On Neuroblastoma Cell Matrix-Related FunctionsdjackapNo ratings yet
- AKT/Protein Kinase B Regulation of BCL Family Members During Oxysterol-Induced ApoptosisDocument8 pagesAKT/Protein Kinase B Regulation of BCL Family Members During Oxysterol-Induced ApoptosisNareshNo ratings yet
- 1 s2.0 S0021925819316795 MainDocument7 pages1 s2.0 S0021925819316795 MainLeidy Constanza Villalobos GonzalezNo ratings yet
- 12 SeminarioDocument6 pages12 SeminarioJonathanNo ratings yet
- Pinealon Increases Cell Viability by Suppression of Free Radical Levels and Activating Proliferative Processes.Document8 pagesPinealon Increases Cell Viability by Suppression of Free Radical Levels and Activating Proliferative Processes.scribdNo ratings yet
- Primary Article 1Document9 pagesPrimary Article 1yusrawasim147No ratings yet
- N6-Methyladenosine Regulates Rna Abundance of Sars-Cov-2: Cell Discovery December 2021Document5 pagesN6-Methyladenosine Regulates Rna Abundance of Sars-Cov-2: Cell Discovery December 2021Ramona AnaNo ratings yet
- In Gel Finger 2015Document10 pagesIn Gel Finger 2015Dumitrache VicentiuNo ratings yet
- 1 FullDocument9 pages1 FullFauziana NurhanisahNo ratings yet
- 9.señalización Osmosensorial en Mycobacterium Tuberculosis 2013Document9 pages9.señalización Osmosensorial en Mycobacterium Tuberculosis 2013Juan Camilo Ocampo MartinezNo ratings yet
- STEM CELLS Translational Med - 2021 - Soto - Neural Crest Like Stem Cells For Tissue RegenerationDocument13 pagesSTEM CELLS Translational Med - 2021 - Soto - Neural Crest Like Stem Cells For Tissue RegenerationAIDAN AsigraNo ratings yet
- Phosphate Regulation of Vascular SmoothDocument9 pagesPhosphate Regulation of Vascular Smoothbryan chanNo ratings yet
- WNT Signaling Multiple PathwaysDocument6 pagesWNT Signaling Multiple PathwaysMayerlis ArrietaNo ratings yet
- Case Report: Rapport de CasDocument3 pagesCase Report: Rapport de CasfadillahidsyamNo ratings yet
- Acute PancreatitisDocument13 pagesAcute PancreatitisJacob BorongNo ratings yet
- Autoantibodies in P45011D6, Cytochrome: LKM-1 Recognize Short Sequence P-450 MonooxygenaseDocument9 pagesAutoantibodies in P45011D6, Cytochrome: LKM-1 Recognize Short Sequence P-450 MonooxygenasefryzinNo ratings yet
- PIIS0021925819643541Document7 pagesPIIS0021925819643541hyouseffardNo ratings yet
- Protein TargetingDocument7 pagesProtein TargetingRaven JohnsonNo ratings yet
- Local Regulation of Extracellular Vesicle Traffic by The Synaptic Endocytic MachineryDocument13 pagesLocal Regulation of Extracellular Vesicle Traffic by The Synaptic Endocytic MachineryValen EstevezNo ratings yet
- Vasopressin ClauserDocument9 pagesVasopressin ClauserwwechiNo ratings yet
- Expression of iNOS in Oral Mucosal Cells by RT-PCR AND ImmunocytochemistryDocument6 pagesExpression of iNOS in Oral Mucosal Cells by RT-PCR AND ImmunocytochemistryrizwanNo ratings yet
- Endothelial Cell NADPH Oxidase Mediates The Cerebral Microvascular Dysfunction in Sickle Cell Transgenic MiceDocument19 pagesEndothelial Cell NADPH Oxidase Mediates The Cerebral Microvascular Dysfunction in Sickle Cell Transgenic MiceNovia MentariNo ratings yet
- Determinants of Postnatal Spleen Tissue Regeneration and OrganogenesisDocument4 pagesDeterminants of Postnatal Spleen Tissue Regeneration and OrganogenesisDaniel Daza BarriosNo ratings yet
- TMP A8 CCDocument13 pagesTMP A8 CCFrontiersNo ratings yet
- Activity 2Document1 pageActivity 2Sebastian CauninNo ratings yet
- Activity 2 2Document1 pageActivity 2 2Sebastian CauninNo ratings yet
- Activity 2 3Document1 pageActivity 2 3Sebastian CauninNo ratings yet
- Activity 3 I. Use The Template or Table Below To Define and Identify The Diseases of The Given Diseases (Picture of The Disease)Document2 pagesActivity 3 I. Use The Template or Table Below To Define and Identify The Diseases of The Given Diseases (Picture of The Disease)Sebastian CauninNo ratings yet
- Is Critical Thinking and Clinical Reasoning The Same? Explain Your AnswerDocument1 pageIs Critical Thinking and Clinical Reasoning The Same? Explain Your AnswerSebastian CauninNo ratings yet
- Act1-PrinciplesofTeaching SebCauninDocument1 pageAct1-PrinciplesofTeaching SebCauninSebastian CauninNo ratings yet
- Protothecosis: Case Report and Literature ReviewDocument12 pagesProtothecosis: Case Report and Literature ReviewSebastian CauninNo ratings yet
- Salary Statement 10 01 2018Document7 pagesSalary Statement 10 01 2018lewin neritNo ratings yet
- Iso 1615 1976Document4 pagesIso 1615 1976Untung HariminNo ratings yet
- BCS - SS-CRM 460 - 2 Jun2019Document2 pagesBCS - SS-CRM 460 - 2 Jun2019Peterson SudlabNo ratings yet
- Departmental Schedule, 2nd Sem 2017-2018Document70 pagesDepartmental Schedule, 2nd Sem 2017-2018Kim Kenneth Roca100% (3)
- Project Plan For Implementation of ISO 20000 20000academy ENDocument7 pagesProject Plan For Implementation of ISO 20000 20000academy ENit20645984 Imashi H.M.G.G.M.No ratings yet
- Copper Pipe Leak Test and VacuumDocument2 pagesCopper Pipe Leak Test and Vacuumalone160162l100% (1)
- Pr2-Chapter3 Mao MN Guto JoDocument5 pagesPr2-Chapter3 Mao MN Guto JoAnaroseNo ratings yet
- Fluxus F/G722: Non-Intrusive Ultrasonic Flow Meter For Highly Dynamic FlowsDocument2 pagesFluxus F/G722: Non-Intrusive Ultrasonic Flow Meter For Highly Dynamic FlowslossaladosNo ratings yet
- First Principle Applications in RoRo-Ship Design PDFDocument7 pagesFirst Principle Applications in RoRo-Ship Design PDFFerdy Fer DNo ratings yet
- IL6 AdvtDocument9 pagesIL6 AdvtManoj Kumar100% (1)
- Digital Signal Processing Exam QuestionsDocument8 pagesDigital Signal Processing Exam QuestionsecehodaietNo ratings yet
- INGLES TECNICO IV (En Ambas Caras)Document45 pagesINGLES TECNICO IV (En Ambas Caras)ESGUAR INSTRUCTORESNo ratings yet
- Cleaning Public Areas EfficientlyDocument231 pagesCleaning Public Areas EfficientlyDj-Ram Vicente Vida100% (6)
- GOLDEN DAWN 3 8 Highlights of The Fourth Knowledge LectureDocument7 pagesGOLDEN DAWN 3 8 Highlights of The Fourth Knowledge LectureF_RCNo ratings yet
- Thefold74 Brooch 1 From Square TubisDocument3 pagesThefold74 Brooch 1 From Square TubisAdriana GranadaNo ratings yet
- XZ100 Partsbook Part2-'09YMDocument60 pagesXZ100 Partsbook Part2-'09YMBikeMaKer BMKNo ratings yet
- How you seize the space between next and nowDocument11 pagesHow you seize the space between next and nowMathan J RNo ratings yet
- Applying The PROACT RCA Methodology.4.26.16Document30 pagesApplying The PROACT RCA Methodology.4.26.16Alexandar ApishaNo ratings yet
- Factors Affecting Online Grocery Shopping Experience: Dhinda Siti Mustikasari, Rifelly Dewi AstutiDocument7 pagesFactors Affecting Online Grocery Shopping Experience: Dhinda Siti Mustikasari, Rifelly Dewi AstutiSiddharth Singh TomarNo ratings yet
- Star Ii HoneywellDocument4 pagesStar Ii HoneywellAlejandro MagnoNo ratings yet
- Modelling of Agglomerating Systems: From Spheres To FractalsDocument13 pagesModelling of Agglomerating Systems: From Spheres To FractalsSandra BazanNo ratings yet
- Human Strategies For A Virtual World: Baldwin BergesDocument32 pagesHuman Strategies For A Virtual World: Baldwin BergesmoisesNo ratings yet
- Ophelia SyndromeDocument10 pagesOphelia SyndromeJulioroncal100% (1)
- User Manual Servo Driver SZGH-302: (One Driver To Control Two Motors Simultaneously)Document23 pagesUser Manual Servo Driver SZGH-302: (One Driver To Control Two Motors Simultaneously)Zdeněk HromadaNo ratings yet
- My ResumeDocument2 pagesMy ResumeSathanandhNo ratings yet
- Evaluation of Green Building in The Existing Building of The Department of The Environment of DKI Jakarta Province in The Order of Reducing Greenhouse Gas EmissionsDocument5 pagesEvaluation of Green Building in The Existing Building of The Department of The Environment of DKI Jakarta Province in The Order of Reducing Greenhouse Gas EmissionsInternational Journal of Innovative Science and Research TechnologyNo ratings yet
- MagnetismDocument30 pagesMagnetismManiratna RoyNo ratings yet
- Sd0001e0 2.0 Con Taratura 957Document37 pagesSd0001e0 2.0 Con Taratura 957JayRellvic Guy-ab0% (1)
- Installation Manual Multi-Function PROTECTION RELAY GRE110, GRE120, GRE130Document22 pagesInstallation Manual Multi-Function PROTECTION RELAY GRE110, GRE120, GRE130Cecep Marfu100% (2)
- VCS Test01Document27 pagesVCS Test01GyanNo ratings yet
- Gut: the new and revised Sunday Times bestsellerFrom EverandGut: the new and revised Sunday Times bestsellerRating: 4 out of 5 stars4/5 (392)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisFrom EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisRating: 3.5 out of 5 stars3.5/5 (2)
- Why We Die: The New Science of Aging and the Quest for ImmortalityFrom EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityRating: 4 out of 5 stars4/5 (3)
- Tales from Both Sides of the Brain: A Life in NeuroscienceFrom EverandTales from Both Sides of the Brain: A Life in NeuroscienceRating: 3 out of 5 stars3/5 (18)
- Fast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperFrom EverandFast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperRating: 4.5 out of 5 stars4.5/5 (15)
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionFrom EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionRating: 4 out of 5 stars4/5 (811)
- The Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorFrom EverandThe Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorNo ratings yet
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessFrom Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessRating: 4 out of 5 stars4/5 (33)
- Gut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)From EverandGut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)Rating: 4 out of 5 stars4/5 (378)
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesFrom EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesRating: 4.5 out of 5 stars4.5/5 (397)
- Who's in Charge?: Free Will and the Science of the BrainFrom EverandWho's in Charge?: Free Will and the Science of the BrainRating: 4 out of 5 stars4/5 (65)
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedFrom EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedRating: 4 out of 5 stars4/5 (11)
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsFrom EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsRating: 4.5 out of 5 stars4.5/5 (4)
- Good Without God: What a Billion Nonreligious People Do BelieveFrom EverandGood Without God: What a Billion Nonreligious People Do BelieveRating: 4 out of 5 stars4/5 (66)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceFrom EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceRating: 4.5 out of 5 stars4.5/5 (515)
- The Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindFrom EverandThe Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindRating: 4.5 out of 5 stars4.5/5 (93)
- Inside of a Dog: What Dogs See, Smell, and KnowFrom EverandInside of a Dog: What Dogs See, Smell, and KnowRating: 4 out of 5 stars4/5 (390)
- Wayfinding: The Science and Mystery of How Humans Navigate the WorldFrom EverandWayfinding: The Science and Mystery of How Humans Navigate the WorldRating: 4.5 out of 5 stars4.5/5 (18)
- This Is Your Brain On Parasites: How Tiny Creatures Manipulate Our Behavior and Shape SocietyFrom EverandThis Is Your Brain On Parasites: How Tiny Creatures Manipulate Our Behavior and Shape SocietyRating: 3.5 out of 5 stars3.5/5 (31)
- Why We Sleep: Unlocking the Power of Sleep and DreamsFrom EverandWhy We Sleep: Unlocking the Power of Sleep and DreamsRating: 4.5 out of 5 stars4.5/5 (2083)
- Crypt: Life, Death and Disease in the Middle Ages and BeyondFrom EverandCrypt: Life, Death and Disease in the Middle Ages and BeyondRating: 4 out of 5 stars4/5 (3)
- A Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouFrom EverandA Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouRating: 4.5 out of 5 stars4.5/5 (62)
- The Dog Who Couldn't Stop Loving: How Dogs Have Captured Our Hearts for Thousands of YearsFrom EverandThe Dog Who Couldn't Stop Loving: How Dogs Have Captured Our Hearts for Thousands of YearsNo ratings yet
- The Invention of Tomorrow: A Natural History of ForesightFrom EverandThe Invention of Tomorrow: A Natural History of ForesightRating: 4.5 out of 5 stars4.5/5 (5)