You might also like
- Biomechanics of GaitDocument27 pagesBiomechanics of Gaitsebastiao_est3197No ratings yet
- Biom Echanical Analysis o F Double Bassists' Perform Ance PosturesDocument164 pagesBiom Echanical Analysis o F Double Bassists' Perform Ance PosturesAlexandre Brasil100% (1)
- Spiro DynamicsDocument12 pagesSpiro DynamicsPooja Chhabra100% (1)
- Biomechanical Analysis of A BallerinaDocument4 pagesBiomechanical Analysis of A BallerinaUrooba ZubairiNo ratings yet
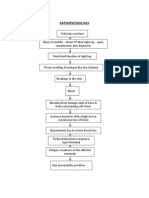
- Fracture PathophysiologyDocument1 pageFracture PathophysiologyIrene Joy Gomez100% (2)
- Human Walking RoseDocument248 pagesHuman Walking RosePablo CruzNo ratings yet
- Jans, M.M.E. Et Al., (2004), 'Characterisation of Microbial Attack On Archaeological Bone', JASDocument9 pagesJans, M.M.E. Et Al., (2004), 'Characterisation of Microbial Attack On Archaeological Bone', JASThomas MafredasNo ratings yet
- Osteop Intl 07Document9 pagesOsteop Intl 07szarysimbaNo ratings yet
- Anatomy and Biomechanics Gluteus Maximus and Thoracolumbar Fascia Sacroiliac JointDocument7 pagesAnatomy and Biomechanics Gluteus Maximus and Thoracolumbar Fascia Sacroiliac JointjoaquinNo ratings yet
- The Human Meniscus: A Review of Anatomy, Function, Injury, and Advances in Treatment: The Meniscus: Anatomy, Function, Injury and TreatmentDocument20 pagesThe Human Meniscus: A Review of Anatomy, Function, Injury, and Advances in Treatment: The Meniscus: Anatomy, Function, Injury and TreatmentLinggaNo ratings yet
- A) The Anatomy in Relation To Injury of The Lateral Collateral Ligaments of The Ankle PDFDocument8 pagesA) The Anatomy in Relation To Injury of The Lateral Collateral Ligaments of The Ankle PDFericdgNo ratings yet
- The Human Meniscus: A Review of Anatomy, Function, Injury, and Advances in Treatment: The Meniscus: Anatomy, Function, Injury and TreatmentDocument20 pagesThe Human Meniscus: A Review of Anatomy, Function, Injury, and Advances in Treatment: The Meniscus: Anatomy, Function, Injury and TreatmentasmaNo ratings yet
- Correlation Compression The Proximal External Degeneration: OF Gross OFDocument10 pagesCorrelation Compression The Proximal External Degeneration: OF Gross OFtrutiNo ratings yet
- The Human Meniscus: A Review of Anatomy, Function, Injury, and Advances in TreatmentDocument19 pagesThe Human Meniscus: A Review of Anatomy, Function, Injury, and Advances in TreatmentRazlivenaTintaNo ratings yet
- O Brien 2005Document6 pagesO Brien 2005prtamaekaputraNo ratings yet
- Comparative Study of Mechanical Stresses in Human Limb BonesDocument9 pagesComparative Study of Mechanical Stresses in Human Limb BonesTejas krishnakanthNo ratings yet
- 1 s2.0 S8756328207006205 MainDocument12 pages1 s2.0 S8756328207006205 Mainhakan sonsuzlukluNo ratings yet
- Lyson Etal 2013 Evolutionary Origin of The Turtle ShellDocument7 pagesLyson Etal 2013 Evolutionary Origin of The Turtle ShellThiago S. SalesNo ratings yet
- Pathways of Load-Induced Cartilage Damage Causing Cartilage Degeneration in The Knee After MeniscectomyDocument7 pagesPathways of Load-Induced Cartilage Damage Causing Cartilage Degeneration in The Knee After MeniscectomyMarcel RodriguezNo ratings yet
- Acumulacion de Microgrietas A Diferentes Intervalos Durante Las Pruebas de Fatiga de Hueso CompactoDocument8 pagesAcumulacion de Microgrietas A Diferentes Intervalos Durante Las Pruebas de Fatiga de Hueso CompactoCamiloVenturaNo ratings yet
- Funtional Morphology of Bite Mechanics in The Great BarracudaDocument14 pagesFuntional Morphology of Bite Mechanics in The Great BarracudaassyahwaNo ratings yet
- Heat-Killed IStaphylococcusDocument14 pagesHeat-Killed IStaphylococcusSilmy IndariNo ratings yet
- Primary Benign and Malignant Osseous Neoplasms of The FootDocument13 pagesPrimary Benign and Malignant Osseous Neoplasms of The FootICNo ratings yet
- The Biomechanics Behind Extreme Osteophagy in Tyrannosaurus RexDocument10 pagesThe Biomechanics Behind Extreme Osteophagy in Tyrannosaurus RexxmirandaNo ratings yet
- Paleoepidemiology, Healing, and Possible Treatment of Trauma in The Medieval Cemetery Population of St. Helen-on-the-Walls, York, EnglandDocument14 pagesPaleoepidemiology, Healing, and Possible Treatment of Trauma in The Medieval Cemetery Population of St. Helen-on-the-Walls, York, EnglandRaluca RainNo ratings yet
- Cotts Et Al. 2019 - Pathology in The Appendicular Bones of Tamandua TetradactylaDocument16 pagesCotts Et Al. 2019 - Pathology in The Appendicular Bones of Tamandua TetradactylaLeonardo CottsNo ratings yet
- McCLEARN, 1985Document29 pagesMcCLEARN, 1985flaviaNo ratings yet
- Tendon Del Subescapular - Sistema de Clasificación - AnatomíaDocument7 pagesTendon Del Subescapular - Sistema de Clasificación - AnatomíajesusglezctNo ratings yet
- Research Article Comparison of Structural, Architectural and Mechanical Aspects of Cellular and Acellular Bone in Two Teleost FishDocument11 pagesResearch Article Comparison of Structural, Architectural and Mechanical Aspects of Cellular and Acellular Bone in Two Teleost FishcamilaNo ratings yet
- Eckstein - Et - Al-2006 - The Effect of Exercise On Human Articular CartilageDocument22 pagesEckstein - Et - Al-2006 - The Effect of Exercise On Human Articular CartilageStavros LitsosNo ratings yet
- Ebacher2007 Dynamic Cimp TractDocument11 pagesEbacher2007 Dynamic Cimp Tractimen mehriNo ratings yet
- Judd One Accident Too ManyDocument13 pagesJudd One Accident Too ManyCristianBeltranAznarezNo ratings yet
- Anderson Et Al. (2012)Document7 pagesAnderson Et Al. (2012)Ruben FernandezNo ratings yet
- 05 - Journal of Anatomy - 2013 - GerlingDocument11 pages05 - Journal of Anatomy - 2013 - Gerlingalan.consultoriafitnessNo ratings yet
- Morphology of The Caudal Vertebrae in RHDocument15 pagesMorphology of The Caudal Vertebrae in RHMIGUEL ANGEL AVILA GUERRANo ratings yet
- Proceedings of The 16th Italian Association of Equine Veterinarians CongressDocument6 pagesProceedings of The 16th Italian Association of Equine Veterinarians CongressCabinet VeterinarNo ratings yet
- 1 s2.0 S0021929012002801 MainDocument6 pages1 s2.0 S0021929012002801 MainCarlos De la cruzNo ratings yet
- A Rare Anomaly of The Clavicle - Bifurcation of The Lateral One-ThirdDocument3 pagesA Rare Anomaly of The Clavicle - Bifurcation of The Lateral One-ThirdSigmundNo ratings yet
- Smith Etal 2006Document6 pagesSmith Etal 2006Jean-Christophe ChampagneNo ratings yet
- Ericpalmarislongus 1Document6 pagesEricpalmarislongus 1CatNo ratings yet
- Bioinspired Design of Exible Armor Based On Chiton Scales: ArticleDocument13 pagesBioinspired Design of Exible Armor Based On Chiton Scales: ArticledragalaurentiuNo ratings yet
- Tingle Et Al, 2023Document20 pagesTingle Et Al, 2023Livia PereiraNo ratings yet
- Barou 2002Document7 pagesBarou 2002Sebastián Hernán Cortés CampillayNo ratings yet
- Osteon: Structure Investigative Applications See Also References Bibliography External LinksDocument3 pagesOsteon: Structure Investigative Applications See Also References Bibliography External LinksAlan FosterNo ratings yet
- Becker 2007Document4 pagesBecker 2007Gustavo RodriguesNo ratings yet
- Limbs in Whales and Limblessness in Other VertebratesDocument14 pagesLimbs in Whales and Limblessness in Other VertebratesJay LeeNo ratings yet
- Diving Adaptations of The Cetacean SkeletonDocument10 pagesDiving Adaptations of The Cetacean SkeletonAngela HurtadoNo ratings yet
- 1602-Article Text-1530-1-1-20190715Document16 pages1602-Article Text-1530-1-1-20190715Mauricio HurtadoNo ratings yet
- Paprosky Classification of Acetabular Bone LossDocument6 pagesPaprosky Classification of Acetabular Bone Lossamine roujdiNo ratings yet
- Form, Function, Food and Feeding in Stellate Echinoderms : ZoosymposiaDocument10 pagesForm, Function, Food and Feeding in Stellate Echinoderms : ZoosymposiaSixto Gutiérrez SaavedraNo ratings yet
- Supination-External Rotation Ankle FracturesDocument9 pagesSupination-External Rotation Ankle FracturesVlad-Andrei SmochinaNo ratings yet
- 7Document3 pages7Deco SoftNo ratings yet
- Fishscale CrackDocument10 pagesFishscale CrackZIJIAN PANNo ratings yet
- Research ArticleDocument5 pagesResearch ArticleZirco HereNo ratings yet
- The Ontogeny of Bite Force Performance IDocument11 pagesThe Ontogeny of Bite Force Performance ICrocodilo VasconcelosNo ratings yet
- The Appendicular Skeleton of Suuwassea eDocument22 pagesThe Appendicular Skeleton of Suuwassea enomadNo ratings yet
- Document 02Document18 pagesDocument 02Marco StagioniNo ratings yet
- Muscle Weakness Causes Joint Degeneration in RabbitsDocument8 pagesMuscle Weakness Causes Joint Degeneration in Rabbitsagonia_00No ratings yet
- Geospiza Bite ForceDocument13 pagesGeospiza Bite ForceHappyMark93No ratings yet
- The Influence of Keel Bone Damage On Welfare of Laying Hens: Anja B. Riber, Teresa M. Casey-Trott and Mette S. HerskinDocument12 pagesThe Influence of Keel Bone Damage On Welfare of Laying Hens: Anja B. Riber, Teresa M. Casey-Trott and Mette S. HerskinPawan MishraNo ratings yet
- Nihms 743965Document25 pagesNihms 743965registrepremsaNo ratings yet
- Structural Design and Mechanical Behavior of Alligator (Alligator Mississippiensis) OsteodermsDocument16 pagesStructural Design and Mechanical Behavior of Alligator (Alligator Mississippiensis) OsteodermsGeorge CatalinNo ratings yet
- Farl Hurlbetal 05 Gator JVP25Document17 pagesFarl Hurlbetal 05 Gator JVP25patelsmita060No ratings yet
- Locomotory Stresses in The Limb Bones of Two Small Mammals: The Ground Squirrel and ChipmunkDocument24 pagesLocomotory Stresses in The Limb Bones of Two Small Mammals: The Ground Squirrel and Chipmunkindjeian111No ratings yet
- Orthodontically Driven Corticotomy: Tissue Engineering to Enhance Orthodontic and Multidisciplinary TreatmentFrom EverandOrthodontically Driven Corticotomy: Tissue Engineering to Enhance Orthodontic and Multidisciplinary TreatmentFederico BrugnamiNo ratings yet
- The Influence of Mechanical Compression On The Induction of Osteoarthritis-Related Biomarkers I N A R T I C U L A R Cartilage ExplantsDocument8 pagesThe Influence of Mechanical Compression On The Induction of Osteoarthritis-Related Biomarkers I N A R T I C U L A R Cartilage ExplantsSTAF OSCENo ratings yet
- Dimitrova Et Al-2012-Principles of Osteoarthritis - Its Definition, Character, Derivation And...Document28 pagesDimitrova Et Al-2012-Principles of Osteoarthritis - Its Definition, Character, Derivation And...STAF OSCENo ratings yet
- The Biomechanics of Osteoarthritis Implications For TherapyDocument8 pagesThe Biomechanics of Osteoarthritis Implications For TherapySTAF OSCENo ratings yet
- Aging and Osteoarthritis The Role of Chondrocyte Senescence and Aging Changes in The Cartilage MatrixDocument9 pagesAging and Osteoarthritis The Role of Chondrocyte Senescence and Aging Changes in The Cartilage MatrixSTAF OSCENo ratings yet
- Ilovepdf Merged 10Document20 pagesIlovepdf Merged 10STAF OSCENo ratings yet
- We Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistsDocument27 pagesWe Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistsSTAF OSCENo ratings yet
- 19 Upaya Peningkatan Kesehatan Persendian Melalui Senam OsteoartritisDocument31 pages19 Upaya Peningkatan Kesehatan Persendian Melalui Senam OsteoartritisSTAF OSCENo ratings yet
- Baseline Articular Contact Stress Tingkats Predict Incident Symptomatic Knee Osteoarthritis Development in The MOST CohortDocument7 pagesBaseline Articular Contact Stress Tingkats Predict Incident Symptomatic Knee Osteoarthritis Development in The MOST CohortSTAF OSCENo ratings yet
- Chondrocyte Hypertrophy and Osteoarthritis Role in Initiation and Progression of Cartilage DegenerationDocument10 pagesChondrocyte Hypertrophy and Osteoarthritis Role in Initiation and Progression of Cartilage DegenerationSTAF OSCENo ratings yet
- The Management of Open Fracture: Gustillo RBDocument21 pagesThe Management of Open Fracture: Gustillo RBsagungw93No ratings yet
- First Aid For Sprain and StrainsDocument9 pagesFirst Aid For Sprain and StrainsKristine May Pangandian MagnoNo ratings yet
- CHAPTER 2 Biomechanics and KinseilogyDocument26 pagesCHAPTER 2 Biomechanics and KinseilogydotrimoNo ratings yet
- Biomechanics and Medical Rehabilitation Lab VedioDocument2 pagesBiomechanics and Medical Rehabilitation Lab VedioTareq KhreisatNo ratings yet
- 2013 Statistical SummaryDocument265 pages2013 Statistical SummaryMarsala Nisto0% (1)
- The Most Prevalent Volleyball Injury Is Patella TendinitisDocument3 pagesThe Most Prevalent Volleyball Injury Is Patella TendinitisKit Jared DolorNo ratings yet
- Fie's Newton LawDocument26 pagesFie's Newton LawNida'ul KhasanahNo ratings yet
- What To Expect: AREA: Music, Arts, and Physical Education (MAPEH) CompetenciesDocument11 pagesWhat To Expect: AREA: Music, Arts, and Physical Education (MAPEH) CompetenciesMarlouNo ratings yet
- Biomechanics of Craniofacial Sutures Orthopedic ImplicationsDocument8 pagesBiomechanics of Craniofacial Sutures Orthopedic ImplicationsMuhammad FahmyNo ratings yet
- Konseling Pada Anak Korban Bencana Alam FixDocument5 pagesKonseling Pada Anak Korban Bencana Alam Fixdeni nasrullahNo ratings yet
- BiomechanicsDocument9 pagesBiomechanicsMuhammad HamzaNo ratings yet
- CAPE PE and Sport SyllabusDocument68 pagesCAPE PE and Sport SyllabusLexi John100% (1)
- Difference Between Sprain and StrainDocument4 pagesDifference Between Sprain and StrainTayyab AliNo ratings yet
- BIE Description of Courses AY1314Document3 pagesBIE Description of Courses AY1314Justin ChongNo ratings yet
- Gait CycleDocument24 pagesGait Cyclebakhtawar safdarNo ratings yet
- Technique Analysis in Sports: A Critical Review: Adrian LeesDocument16 pagesTechnique Analysis in Sports: A Critical Review: Adrian LeesRuy ZuluNo ratings yet
- Thoracic TraumaDocument12 pagesThoracic TraumaNiarti Ulan Sari SiarnuNo ratings yet
- Jamia Milia Islamia MPT Syllabus PDFDocument118 pagesJamia Milia Islamia MPT Syllabus PDFMrs. SHARADA LAKSHMANA NAYAK AI001901No ratings yet
- Injury Report Form: Circle and NameDocument1 pageInjury Report Form: Circle and NameNinNo ratings yet
- Proceedings WCCM 2014Document37 pagesProceedings WCCM 2014HectorNo ratings yet
- SPS Week 3Document6 pagesSPS Week 3aimeeNo ratings yet
- Tchernik Ioulia 2017 MADocument130 pagesTchernik Ioulia 2017 MAsergio leoneNo ratings yet
- SearchOok Results For MechanicDocument30 pagesSearchOok Results For Mechaniclche0590% (1)