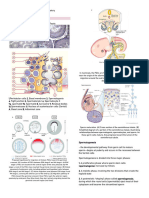
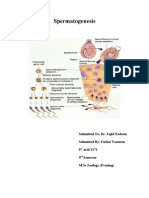
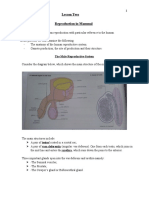
You might also like
- 11 MaleDocument14 pages11 MaleJasmine KaurNo ratings yet
- SPERMATOGENESISDocument58 pagesSPERMATOGENESISkathyNo ratings yet
- CC-13 Unit 1Document10 pagesCC-13 Unit 1Subhradeep SinghaNo ratings yet
- SPERMATOGENESIS The First Wave of Spermatogenesis Moreno 2018Document6 pagesSPERMATOGENESIS The First Wave of Spermatogenesis Moreno 2018LudimilaNo ratings yet
- Devl Biol-SpermatogenesisDocument9 pagesDevl Biol-SpermatogenesisSuresh DavuluriNo ratings yet
- Biology Notes: Key PointsDocument15 pagesBiology Notes: Key PointsAngelineBagtasNo ratings yet
- Hormonal Control of GametogenesisDocument29 pagesHormonal Control of GametogenesisWachiel Arhamz100% (2)
- Gametogenesis Process in HumanDocument10 pagesGametogenesis Process in HumanFerhaeeza KalayakanNo ratings yet
- Spermatogenesis WPS Office 1Document12 pagesSpermatogenesis WPS Office 1nb6pd5rvnxNo ratings yet
- Histology of The Male Reproductive SystemDocument18 pagesHistology of The Male Reproductive SystemMuhammad JameelNo ratings yet
- Spermatogenesis:: (1) Formation of TheDocument3 pagesSpermatogenesis:: (1) Formation of ThebarbacumlaudeNo ratings yet
- Spermatogenesis Is The Process by Which: Seminiferous Tubule With Maturing Sperm.Document2 pagesSpermatogenesis Is The Process by Which: Seminiferous Tubule With Maturing Sperm.Garcia AzirNo ratings yet
- GametogenesisDocument3 pagesGametogenesisWai KikiNo ratings yet
- Cell Division: There Are Two Types of Cell Divisions Which Are andDocument23 pagesCell Division: There Are Two Types of Cell Divisions Which Are andapi-3706483No ratings yet
- Anbt - 608: Submitted ToDocument50 pagesAnbt - 608: Submitted ToMayuriGulhaneNo ratings yet
- Spermatogenesis: Feriha Ercan, Ph.D. Dept. Histology and EmbryologyDocument41 pagesSpermatogenesis: Feriha Ercan, Ph.D. Dept. Histology and EmbryologyErdem AltunNo ratings yet
- LAB EXERCISE 1 - SpermatogenesisDocument5 pagesLAB EXERCISE 1 - SpermatogenesisGerald Angelo DeguinioNo ratings yet
- Developmental Biology Notes 3Document26 pagesDevelopmental Biology Notes 3PrezyNo ratings yet
- 1999 Ency HessDocument8 pages1999 Ency HessKishan GaurNo ratings yet
- Department of Morphology: Penza State University Medical InstituteDocument18 pagesDepartment of Morphology: Penza State University Medical InstituteunnatiNo ratings yet
- The Human Reproductive SystemDocument7 pagesThe Human Reproductive SystemAlloiza CaguiclaNo ratings yet
- S H I. II. S O, M Iii. I IV. S - G, M, E, P, T I. II. Iii. IV. V. VI. ViiDocument6 pagesS H I. II. S O, M Iii. I IV. S - G, M, E, P, T I. II. Iii. IV. V. VI. ViistriataNo ratings yet
- Gametogenesis: Gametogenesis Is The Process by Which Diploid Precursor Cells Undergo MeioticDocument2 pagesGametogenesis: Gametogenesis Is The Process by Which Diploid Precursor Cells Undergo MeioticnolaNo ratings yet
- 2B Dev BioDocument63 pages2B Dev BioYashNo ratings yet
- DR Anangwe-GametogenesisDocument23 pagesDR Anangwe-GametogenesisStanley OdiraNo ratings yet
- Introduction To PhysiologyDocument9 pagesIntroduction To PhysiologyTimothy usmanNo ratings yet
- Gametogenesis Process in HumanDocument13 pagesGametogenesis Process in HumanRiski UntariNo ratings yet
- Development of Fish Oocytes During ChromatinDocument5 pagesDevelopment of Fish Oocytes During ChromatinCristel Mae De GuzmanNo ratings yet
- Regeneration in AxolotlDocument13 pagesRegeneration in AxolotlȺនђᎥនђ ツᦔøץNo ratings yet
- Gametogenesis, Gamete Transport & FertilizationDocument56 pagesGametogenesis, Gamete Transport & FertilizationvictorNo ratings yet
- 11.4 Sexual Reproduction: Name .ClassDocument21 pages11.4 Sexual Reproduction: Name .ClassReshma DookarranNo ratings yet
- SPERMATOGENESISDocument9 pagesSPERMATOGENESISJustine Ian Sean CorpuzNo ratings yet
- EMBRIOLOGYDocument36 pagesEMBRIOLOGYHelena Pascual PérezNo ratings yet
- Physiology II - PresentationDocument11 pagesPhysiology II - PresentationalshmryalmwydNo ratings yet
- GametogenesisDocument35 pagesGametogenesisJason TulipatNo ratings yet
- Embryo PDFDocument80 pagesEmbryo PDFMOHSEN PMNo ratings yet
- GametogenesisDocument8 pagesGametogenesisSOPHIA ALESNANo ratings yet
- 11.4 - Reproduction IB BiologyDocument3 pages11.4 - Reproduction IB BiologyannehindenbergNo ratings yet
- SpermatogenesisDocument16 pagesSpermatogenesisFazilaNo ratings yet
- Q & A Notebook Chap 03Document13 pagesQ & A Notebook Chap 03umamahfarooq75No ratings yet
- GametogenesisDocument5 pagesGametogenesisjabreen ahmadNo ratings yet
- Male Reproductive SystemDocument28 pagesMale Reproductive SystemGenerato, Todd MatthewNo ratings yet
- SpermatogenesisDocument8 pagesSpermatogenesisapi-26413035No ratings yet
- Module 3 CytogeneticsDocument23 pagesModule 3 CytogeneticsFrances Riane SimoyNo ratings yet
- Plasmodium Vivax Malarial ParasiteDocument32 pagesPlasmodium Vivax Malarial Parasiteyuvrajchaudhary982No ratings yet
- Biology Investigatory Project On GametogenesisDocument20 pagesBiology Investigatory Project On GametogenesisAshwin Rockzz46% (13)
- Spermatogenesis: Created by Aji SantikoDocument16 pagesSpermatogenesis: Created by Aji SantikoPrince of Darkness100% (1)
- General Embryology: Gametogenesis: Formation of Male & Female GametesDocument19 pagesGeneral Embryology: Gametogenesis: Formation of Male & Female Gametes이희승No ratings yet
- Objective:-To Study Gametogensis in Human BeingDocument8 pagesObjective:-To Study Gametogensis in Human BeingKUNAL1221No ratings yet
- M SC Zoology III Sem Paper II DR S K ThakurDocument6 pagesM SC Zoology III Sem Paper II DR S K ThakurAnisha SalujaNo ratings yet
- Intoduction To Embryology, Testis & Spermatogenesis - Nto, Nto JohnsonDocument26 pagesIntoduction To Embryology, Testis & Spermatogenesis - Nto, Nto JohnsonigboegwummesomaNo ratings yet
- Lesson Two - Reproduction in MammalsDocument5 pagesLesson Two - Reproduction in MammalsNobody JamesNo ratings yet
- Spermatogenesis - Biology Science Fair Project IdeasDocument8 pagesSpermatogenesis - Biology Science Fair Project IdeasEPFO AKURDI100% (1)
- Chapter 28 - The Reproductive SystemDocument7 pagesChapter 28 - The Reproductive Systemtomorrow.today.yesterday .yesterdayNo ratings yet
- Neiron GametogenesisDocument6 pagesNeiron GametogenesisJose MartinezNo ratings yet
- SpermatogenesisDocument13 pagesSpermatogenesiszuha khanNo ratings yet
- Development of AmphioxusDocument59 pagesDevelopment of AmphioxusDyah Ayu PuspitasariNo ratings yet
- Importance of Human Reproductive SystemDocument3 pagesImportance of Human Reproductive Systemshn mzkNo ratings yet
- The Male Genital SystemDocument5 pagesThe Male Genital SystemDr-AymanBahgatNo ratings yet
- Rhumatologie-Pediatrique FRDocument4 pagesRhumatologie-Pediatrique FRCaity YoungNo ratings yet
- Hypoxic Ischaemic Encephalopathy HIE AsphyxiaDocument6 pagesHypoxic Ischaemic Encephalopathy HIE AsphyxiaCaity YoungNo ratings yet
- Afoem Sample Paper Stage B Written Exam Paper 7Document46 pagesAfoem Sample Paper Stage B Written Exam Paper 7Caity YoungNo ratings yet
- A I - E M: Drenal Nsufficiency Mergency AnagementDocument12 pagesA I - E M: Drenal Nsufficiency Mergency AnagementCaity YoungNo ratings yet
- Fluid Calculator DKADocument3 pagesFluid Calculator DKACaity YoungNo ratings yet
- Afoem Sample Paper Stage B Written Exam Paper 7Document43 pagesAfoem Sample Paper Stage B Written Exam Paper 7Caity YoungNo ratings yet
- 2014 A Health Professionals Guide To Using The ChartsDocument14 pages2014 A Health Professionals Guide To Using The ChartsCaity YoungNo ratings yet
- Correspondence: Classifi Cation of CFTR Mutation ClassesDocument2 pagesCorrespondence: Classifi Cation of CFTR Mutation ClassesCaity YoungNo ratings yet
- SSHADESS HandoutDocument1 pageSSHADESS HandoutCaity YoungNo ratings yet
- Reference Ranges For Urinalysis - Microbiology: Test Range Units DateDocument1 pageReference Ranges For Urinalysis - Microbiology: Test Range Units DateCaity YoungNo ratings yet
- ToF ScriptDocument6 pagesToF ScriptCaity YoungNo ratings yet
- 2 - McDonald - Examples - of - Unhelpful - ThoughtsDocument2 pages2 - McDonald - Examples - of - Unhelpful - ThoughtsCaity YoungNo ratings yet
- Genetic Basis For Congenital Heart Disease: Revisited: CirculationDocument59 pagesGenetic Basis For Congenital Heart Disease: Revisited: CirculationCaity YoungNo ratings yet
- 4-5 GlomerularDiseasesDocument21 pages4-5 GlomerularDiseasesCaity YoungNo ratings yet
- IsbelDocument7 pagesIsbelCaity YoungNo ratings yet
- Sore Throat: Navigating The Differential DiagnosisDocument7 pagesSore Throat: Navigating The Differential DiagnosisCaity YoungNo ratings yet
- During and at The End of The 60-Minute Discussion, The Learners Should Be Able ToDocument6 pagesDuring and at The End of The 60-Minute Discussion, The Learners Should Be Able ToKathNo ratings yet
- Human Reproductive SystemDocument8 pagesHuman Reproductive SystemDivyanshu AggarwalNo ratings yet
- General Biology 1 - Module 7-MitosisandmeiosisDocument32 pagesGeneral Biology 1 - Module 7-Mitosisandmeiosisエアーア ラシブNo ratings yet
- Bio IC Lesson 7 - MutationDocument13 pagesBio IC Lesson 7 - MutationKurt KleinNo ratings yet
- ChlamydomonasDocument9 pagesChlamydomonasPunam TamangNo ratings yet
- Human Anatomy & Physiology I - Dr. Sullivan Unit IV - Cellular Function Chapter 4, Chapter 27 (Meiosis Only)Document5 pagesHuman Anatomy & Physiology I - Dr. Sullivan Unit IV - Cellular Function Chapter 4, Chapter 27 (Meiosis Only)Steve SullivanNo ratings yet
- Kebo 103Document14 pagesKebo 103Cedrickk AlforooNo ratings yet
- GENBIO 1 Q1 Periodic TestDocument6 pagesGENBIO 1 Q1 Periodic TestCelsa ChicoNo ratings yet
- Science 8 Quarter 4 Shortened Module 2 CELL DIVISION Week 2Document6 pagesScience 8 Quarter 4 Shortened Module 2 CELL DIVISION Week 2Alvin OliverosNo ratings yet
- 10 Cell Cycle N Cell Division-NotesDocument4 pages10 Cell Cycle N Cell Division-NotesAnanth Dharanidharan100% (1)
- Biology: Performance Enhancement Sheet - 81Document1 pageBiology: Performance Enhancement Sheet - 81Jai Mata DiNo ratings yet
- Serial-Position EffectDocument6 pagesSerial-Position Effectisbro1788No ratings yet
- Activity 18 AgingDocument27 pagesActivity 18 AgingKendrix Aguiluz FlorendoNo ratings yet
- NCERT Solutions For Class 12 March 30 Biology Chapter 1 Reproduction in OrganismsDocument6 pagesNCERT Solutions For Class 12 March 30 Biology Chapter 1 Reproduction in Organismstechno indiaNo ratings yet
- Identification of General Transcription Factor-Two-E and Mediator Complex Interactions in The Context of Pre-Initiation Complex FormationDocument83 pagesIdentification of General Transcription Factor-Two-E and Mediator Complex Interactions in The Context of Pre-Initiation Complex Formationmunevver099kandemirNo ratings yet
- Mutations WSDocument3 pagesMutations WSNixoniaNo ratings yet
- Cytological Bases of HeredityDocument42 pagesCytological Bases of Heredity'tephanie Salcedo100% (1)
- Genetica Humana BetaDocument70 pagesGenetica Humana BetamiltonlafebreNo ratings yet
- Cardiac RegenerationDocument5 pagesCardiac RegenerationBenyam ZenebeNo ratings yet
- Genomic Equivalence: DefinitionDocument5 pagesGenomic Equivalence: Definitionjgfjhf arwtrNo ratings yet
- F21 - BIOL 1020 Lab 5 HandoutDocument11 pagesF21 - BIOL 1020 Lab 5 HandoutNoyaNo ratings yet
- Mutation AssignmentDocument1 pageMutation AssignmentANNE CZARINA RAMIELLE DE VILLANo ratings yet
- PlasmoDocument1 pagePlasmonajanunhoikiasilNo ratings yet
- Plant Kingdom Class 11 Notes NEET Biology (PDF)Document17 pagesPlant Kingdom Class 11 Notes NEET Biology (PDF)shusa harshaNo ratings yet
- 2015 Molecular Biology of The Cell 6th Edition Test BankDocument29 pages2015 Molecular Biology of The Cell 6th Edition Test Bankpeggymccallxnpsogirke100% (39)
- Polyspermy and How To Reduce ItDocument35 pagesPolyspermy and How To Reduce ItbrachyotisNo ratings yet
- Prepared By: K.Y.LOHDocument26 pagesPrepared By: K.Y.LOHNirmell SatthiyasilanNo ratings yet
- Monday Tuesday Wednesday Thursday Friday: GRADES 1 To 12 Daily Lesson LogDocument9 pagesMonday Tuesday Wednesday Thursday Friday: GRADES 1 To 12 Daily Lesson LogJerwin Monsales100% (1)
- 05 - Growth & Reproduction Exercise 3Document2 pages05 - Growth & Reproduction Exercise 3rhdoiuaNo ratings yet
- Cell Proliferation, Apoptosis, Repair and Regeneration - ClinicalKeyDocument25 pagesCell Proliferation, Apoptosis, Repair and Regeneration - ClinicalKeyMd Jahidul IslamNo ratings yet
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessFrom Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessRating: 4 out of 5 stars4/5 (33)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceFrom EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceRating: 4.5 out of 5 stars4.5/5 (517)
- Why We Die: The New Science of Aging and the Quest for ImmortalityFrom EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityRating: 4 out of 5 stars4/5 (5)
- Tales from Both Sides of the Brain: A Life in NeuroscienceFrom EverandTales from Both Sides of the Brain: A Life in NeuroscienceRating: 3 out of 5 stars3/5 (18)
- Return of the God Hypothesis: Three Scientific Discoveries That Reveal the Mind Behind the UniverseFrom EverandReturn of the God Hypothesis: Three Scientific Discoveries That Reveal the Mind Behind the UniverseRating: 4.5 out of 5 stars4.5/5 (52)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisFrom EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisRating: 3.5 out of 5 stars3.5/5 (2)
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedFrom EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedRating: 4 out of 5 stars4/5 (11)
- Gut: the new and revised Sunday Times bestsellerFrom EverandGut: the new and revised Sunday Times bestsellerRating: 4 out of 5 stars4/5 (393)
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionFrom EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionRating: 4 out of 5 stars4/5 (812)
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesFrom EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesRating: 4.5 out of 5 stars4.5/5 (397)
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsFrom EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsRating: 4.5 out of 5 stars4.5/5 (6)
- Good Without God: What a Billion Nonreligious People Do BelieveFrom EverandGood Without God: What a Billion Nonreligious People Do BelieveRating: 4 out of 5 stars4/5 (66)
- Seven and a Half Lessons About the BrainFrom EverandSeven and a Half Lessons About the BrainRating: 4 out of 5 stars4/5 (110)
- Buddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomFrom EverandBuddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomRating: 4 out of 5 stars4/5 (216)
- Who's in Charge?: Free Will and the Science of the BrainFrom EverandWho's in Charge?: Free Will and the Science of the BrainRating: 4 out of 5 stars4/5 (65)
- The Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorFrom EverandThe Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorNo ratings yet
- The Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindFrom EverandThe Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindRating: 4.5 out of 5 stars4.5/5 (93)
- Gut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)From EverandGut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)Rating: 4 out of 5 stars4/5 (411)
- The Lives of Bees: The Untold Story of the Honey Bee in the WildFrom EverandThe Lives of Bees: The Untold Story of the Honey Bee in the WildRating: 4.5 out of 5 stars4.5/5 (44)
- Darwin's Doubt: The Explosive Origin of Animal Life and the Case for Intelligent DesignFrom EverandDarwin's Doubt: The Explosive Origin of Animal Life and the Case for Intelligent DesignRating: 4.5 out of 5 stars4.5/5 (39)
- The Rise and Fall of the Dinosaurs: A New History of a Lost WorldFrom EverandThe Rise and Fall of the Dinosaurs: A New History of a Lost WorldRating: 4 out of 5 stars4/5 (596)
- Moral Tribes: Emotion, Reason, and the Gap Between Us and ThemFrom EverandMoral Tribes: Emotion, Reason, and the Gap Between Us and ThemRating: 4.5 out of 5 stars4.5/5 (115)
- Human: The Science Behind What Makes Your Brain UniqueFrom EverandHuman: The Science Behind What Makes Your Brain UniqueRating: 3.5 out of 5 stars3.5/5 (38)
- Fast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperFrom EverandFast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperRating: 4.5 out of 5 stars4.5/5 (16)