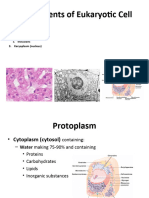
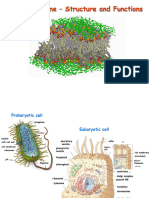
You might also like
- A-level Biology Revision: Cheeky Revision ShortcutsFrom EverandA-level Biology Revision: Cheeky Revision ShortcutsRating: 5 out of 5 stars5/5 (5)
- Membranes 10 31 2012 NotesDocument18 pagesMembranes 10 31 2012 Notesapi-178321548No ratings yet
- Membranes and Proteins: Structure and FunctionDocument52 pagesMembranes and Proteins: Structure and FunctionBlaNo ratings yet
- Pale Child with Splenomegaly and Hemolytic AnemiaDocument49 pagesPale Child with Splenomegaly and Hemolytic AnemiaAwais RehmanNo ratings yet
- 8-Plasma Membranes 28-7-2020Document37 pages8-Plasma Membranes 28-7-2020Hiba N IkhmyesNo ratings yet
- Cell Structure and Function For 1st Year MBBS Delivered by Miss Sumaira On 25 Jan 2010Document61 pagesCell Structure and Function For 1st Year MBBS Delivered by Miss Sumaira On 25 Jan 2010IMDCBiochem83% (6)
- Plasma Membrane: The Lipid Bilayer Membrane ProteinsDocument79 pagesPlasma Membrane: The Lipid Bilayer Membrane ProteinsEarfanNo ratings yet
- Biological MembranesDocument8 pagesBiological MembranesT4No ratings yet
- 1.2 Cell PhysiologyDocument39 pages1.2 Cell PhysiologyChristone “Zuluzulu” ZuluNo ratings yet
- 4.cell 1Document67 pages4.cell 1Raymond LundaNo ratings yet
- The Cell Structure, Functions of The CellDocument76 pagesThe Cell Structure, Functions of The CellCharles SsekawuNo ratings yet
- Membrane Structure and Function: A Fluid Mosaic ModelDocument15 pagesMembrane Structure and Function: A Fluid Mosaic ModeldndqdqNo ratings yet
- MembranesDocument50 pagesMembranesStanley ChikoveNo ratings yet
- L5 Transport MechanismsDocument34 pagesL5 Transport MechanismsAlghester PiangcoNo ratings yet
- Biol2120 Cell Biology: Membranes: Their Structure, Function, and Chemistry 2. MembranesDocument49 pagesBiol2120 Cell Biology: Membranes: Their Structure, Function, and Chemistry 2. MembranesHui Ka HoNo ratings yet
- Plant Cell Structure and FunctionDocument29 pagesPlant Cell Structure and Functionihebunnaogochukwu24No ratings yet
- Cell Structure and PhysiologyDocument29 pagesCell Structure and PhysiologyZaid AbdulqadirNo ratings yet
- AP BIO MACROMOLECULES Chapter 5 ReviewDocument35 pagesAP BIO MACROMOLECULES Chapter 5 Reviewsheila may valiao-de asisNo ratings yet
- Structure and Function of MacromoleculesDocument35 pagesStructure and Function of Macromoleculesrina muliawanNo ratings yet
- The Basic Unit of Life - The Cell and Its FunctionsDocument100 pagesThe Basic Unit of Life - The Cell and Its FunctionsDell E6430No ratings yet
- Bio MoleculesDocument47 pagesBio MoleculesVivaMapwaNo ratings yet
- Cell MembraneDocument32 pagesCell Membraneour lectureNo ratings yet
- 1 Lec.1 (w1) Overview and Plant StructureDocument32 pages1 Lec.1 (w1) Overview and Plant StructureRabiatul AdawiyyahNo ratings yet
- A407433177 - 23745 - 23 - 2019 - Plasma Membrane - Compressed PDFDocument111 pagesA407433177 - 23745 - 23 - 2019 - Plasma Membrane - Compressed PDFDiotima BhattacharyaNo ratings yet
- Structure of Cell Membrane & Cellular Junctions: Pradeep Singh M.Sc. Medical Biochemistry Himsr, Jamia HamdardDocument37 pagesStructure of Cell Membrane & Cellular Junctions: Pradeep Singh M.Sc. Medical Biochemistry Himsr, Jamia HamdardFarhan AhmedNo ratings yet
- L2 Cell Membrane and Transport 1Document31 pagesL2 Cell Membrane and Transport 1sampsonsoo17No ratings yet
- 2 Cell Structure - A 2019Document88 pages2 Cell Structure - A 2019Isworo RukmiNo ratings yet
- ProteinsDocument59 pagesProteinsCraig MitchellNo ratings yet
- Plasma Membrane: Created By: Dyah Candra Palupi Syarifatul Laili Anggun Arifka Diah Ajeng SDocument36 pagesPlasma Membrane: Created By: Dyah Candra Palupi Syarifatul Laili Anggun Arifka Diah Ajeng SDwiRaniPrihandiniNo ratings yet
- The Cell: and Cellular OrganellesDocument70 pagesThe Cell: and Cellular OrganellesKunda JosephNo ratings yet
- CELL BIOLOGY TITLEDocument75 pagesCELL BIOLOGY TITLERhea SharmaNo ratings yet
- Chapter 4 Cell Membranes and TransportDocument33 pagesChapter 4 Cell Membranes and TransportChia Hon LamNo ratings yet
- Prokaryotic and Eukaryotic CellsDocument78 pagesProkaryotic and Eukaryotic CellsLandon GlascockNo ratings yet
- Cell PhysiologyDocument45 pagesCell PhysiologyHimani JhaNo ratings yet
- Biological MembranesDocument31 pagesBiological MembranesHimani Jha100% (1)
- Physiology Chapter 2 Part 2-1Document42 pagesPhysiology Chapter 2 Part 2-1prabokrishna02No ratings yet
- Cell - Basic Unit of Life.Document14 pagesCell - Basic Unit of Life.Ayesha MasoodNo ratings yet
- Cell PhysiologyDocument40 pagesCell PhysiologyAmmara KhalidNo ratings yet
- Biology Unit Chapters 2 and 3 Plus IBDocument164 pagesBiology Unit Chapters 2 and 3 Plus IBallie3d100% (1)
- The Cell Membrane and OrganellesDocument16 pagesThe Cell Membrane and OrganellesChris GayleNo ratings yet
- Introduction To BiochemistryDocument55 pagesIntroduction To BiochemistryHamzaNo ratings yet
- Hist of Cell-Organelles 2022Document45 pagesHist of Cell-Organelles 2022betelhema71No ratings yet
- Pengantar Biologi SelDocument81 pagesPengantar Biologi SeldiesNo ratings yet
- Cell MembraneDocument100 pagesCell MembranephyMedza AppNo ratings yet
- Membrane ProteinsDocument47 pagesMembrane ProteinsStanley ChikoveNo ratings yet
- L03 Membrane StructureDocument37 pagesL03 Membrane StructureMa Christina Alessandra HingcoNo ratings yet
- Cellular Macromolecules and Their FunctionsDocument40 pagesCellular Macromolecules and Their FunctionsidreesnazimNo ratings yet
- Cell Structure & Function 2022Document43 pagesCell Structure & Function 2022Kavita MahaseNo ratings yet
- Cell MembraneDocument16 pagesCell Membraneeishawajid75No ratings yet
- Cells & Physiology - Lecture (Week 3-4)Document59 pagesCells & Physiology - Lecture (Week 3-4)Trevannie EdwardsNo ratings yet
- Basics of Human CellDocument24 pagesBasics of Human Cellananya girishbabuNo ratings yet
- Chapter 5Document60 pagesChapter 5EhazNo ratings yet
- Chapter 7 by Hoch PDFDocument51 pagesChapter 7 by Hoch PDFWynterNo ratings yet
- Plasma Membrane and Transport MechanismsDocument63 pagesPlasma Membrane and Transport MechanismsVaccedNo ratings yet
- Struktur SelDocument82 pagesStruktur SelIda Fitria RahmawatiNo ratings yet
- Gen - Bio Week 1Document62 pagesGen - Bio Week 1yahnie ًNo ratings yet
- Cell DR Fasiha Hussain Biochemistry DemonstratorDocument37 pagesCell DR Fasiha Hussain Biochemistry DemonstratorAqsa ArifNo ratings yet
- Chapter 3-2Document62 pagesChapter 3-2Sifan MotumaNo ratings yet
- Tissue (2021 - 01 - 21 00 - 20 - 54 UTC)Document33 pagesTissue (2021 - 01 - 21 00 - 20 - 54 UTC)Shalik WalkerNo ratings yet
- Biology I For Non-Majors: Module 3: Important Biological MacromoleculesDocument17 pagesBiology I For Non-Majors: Module 3: Important Biological MacromoleculesEmma RiftyanNo ratings yet
- Cell Membrane AA 2023Document40 pagesCell Membrane AA 2023Hurmat AmnaNo ratings yet
- Membrane Structure and FunctionDocument46 pagesMembrane Structure and FunctionHurmat AmnaNo ratings yet
- ch10 Biomembrane structure-LODISHDocument79 pagesch10 Biomembrane structure-LODISHHurmat AmnaNo ratings yet
- Assignment 1 Hurmat e Amna 2020 Ag 8712Document10 pagesAssignment 1 Hurmat e Amna 2020 Ag 8712Hurmat AmnaNo ratings yet
- Crystal MorphologyDocument41 pagesCrystal Morphologyagvega69109No ratings yet
- Biochemistry ReviewDocument13 pagesBiochemistry ReviewMohammad IzadiNo ratings yet
- 1 EBSD PrincipleDocument49 pages1 EBSD PrincipleandrianidebrinaNo ratings yet
- Antigen Presentation SlidesDocument53 pagesAntigen Presentation SlidesShakti PatelNo ratings yet
- Biology NotesDocument3 pagesBiology Notespenelope stieglitzNo ratings yet
- 5year Int. M.sc. in CMB, 2014 Admission BroucherDocument7 pages5year Int. M.sc. in CMB, 2014 Admission BroucherPrashant PatelNo ratings yet
- Chapter 3 - Cells & TissuesDocument197 pagesChapter 3 - Cells & TissuesSean Vladimir SorianoNo ratings yet
- 1 Cells of The Immune SystemDocument90 pages1 Cells of The Immune Systemmark njeru ngigiNo ratings yet
- Revised XRD Experiment ManualDocument9 pagesRevised XRD Experiment ManualAina SyaqinaNo ratings yet
- Chapter 4 Chemical Composition of The CellDocument9 pagesChapter 4 Chemical Composition of The CellWONG LI KING MoeNo ratings yet
- Miller Indices for Planes in Cubic Unit CellsDocument2 pagesMiller Indices for Planes in Cubic Unit CellsAndree Scepter Guansing100% (1)
- Question 1: Identify On A Cell Model or Diagram The Three Major Cell Regions (NucleusDocument10 pagesQuestion 1: Identify On A Cell Model or Diagram The Three Major Cell Regions (NucleusGracea Mae AranetaNo ratings yet
- Of Solids: PolymorphismDocument18 pagesOf Solids: PolymorphismShantanu PaulNo ratings yet
- Set A Suggested Answer Mosys SB025 - 230430 - 191516Document4 pagesSet A Suggested Answer Mosys SB025 - 230430 - 191516nursyakiramailanNo ratings yet
- Biology ProjectDocument6 pagesBiology ProjectBemnet TayeNo ratings yet
- Barsanalina - Assignment For Chapter 16 Adaptive ImmunityDocument4 pagesBarsanalina - Assignment For Chapter 16 Adaptive ImmunityRalph B. BarsanalinaNo ratings yet
- Protein - Structures and FunctionsDocument33 pagesProtein - Structures and FunctionsGracentya AninNo ratings yet
- Unit 2: Cell Structure and Function Chapters 6-7Document4 pagesUnit 2: Cell Structure and Function Chapters 6-7api-542216138No ratings yet
- Chapter - 03 - 2-Crystal StructuresDocument24 pagesChapter - 03 - 2-Crystal Structuresyelten14No ratings yet
- LSE01-B2-Transport Molecules and Proton PumpsDocument15 pagesLSE01-B2-Transport Molecules and Proton PumpsAD-MQNo ratings yet
- Chemiosmosis ExplainedDocument4 pagesChemiosmosis Explainedramloghun veer100% (1)
- Introduction and Overview of Immune SystemDocument14 pagesIntroduction and Overview of Immune SystemEssington BeloNo ratings yet
- Introduction To Materials Science and Engineering: Lecture-3Document14 pagesIntroduction To Materials Science and Engineering: Lecture-3Sibasish RoutNo ratings yet
- Antigen-Presenting CellsDocument9 pagesAntigen-Presenting Cellsnur fitrianaNo ratings yet
- NMR SpectrosDocument1 pageNMR SpectrosFaiza SibtainNo ratings yet
- Carbohydrates, Lipids, Proteins TableDocument1 pageCarbohydrates, Lipids, Proteins TableCarmela VargasNo ratings yet
- Introduction To Biochemistry: by Zaheer Uddin (Pharm-D, M Phil (Pharmacy Practice) MBA (MARKETING)Document63 pagesIntroduction To Biochemistry: by Zaheer Uddin (Pharm-D, M Phil (Pharmacy Practice) MBA (MARKETING)Izat KhanNo ratings yet
- Introduction To BiophysicsDocument9 pagesIntroduction To BiophysicszhairaffeNo ratings yet
- Solution Manual For Materials For Civil and Construction Engineers 4Th Edition Mamlouk Zaniewski 0134320530 9780134320533 Full Chapter PDFDocument28 pagesSolution Manual For Materials For Civil and Construction Engineers 4Th Edition Mamlouk Zaniewski 0134320530 9780134320533 Full Chapter PDFrolland.pletcher494100% (11)
- Movement of Substances Across The Plasma Membrane: Compiled & Prepared by Dsalleh (2021)Document25 pagesMovement of Substances Across The Plasma Membrane: Compiled & Prepared by Dsalleh (2021)Dewi SallehNo ratings yet