You might also like
- Genetics in OrthodonticsDocument63 pagesGenetics in OrthodonticsPrachi Madan100% (20)
- Single Gene Inheritance HandoutDocument16 pagesSingle Gene Inheritance HandoutMarel LatawanNo ratings yet
- Edmonson - Pageantry Overture - AnalysisDocument3 pagesEdmonson - Pageantry Overture - Analysisapi-426112870No ratings yet
- Molecular Biology: An Introduction and BackgroundDocument22 pagesMolecular Biology: An Introduction and Backgroundsyedamaryam zahidNo ratings yet
- Genetics Learning Materials Bsa 2 1Document34 pagesGenetics Learning Materials Bsa 2 1Khy Renz Bajaj100% (1)
- 1st Lecture History of Molecular BiologyDocument52 pages1st Lecture History of Molecular BiologyShanza SyedNo ratings yet
- Patterns of InheritanceDocument47 pagesPatterns of InheritanceFrenche PañaresNo ratings yet
- Genetics: From Mendel to DNADocument5 pagesGenetics: From Mendel to DNAYassine OUEDRAOGONo ratings yet
- 1 Week Lecture1-2 (Credit Hour 1,2)Document4 pages1 Week Lecture1-2 (Credit Hour 1,2)Айжан МубараковаNo ratings yet
- Genetics lesson 2 assessmentDocument10 pagesGenetics lesson 2 assessmentRichelle MasingNo ratings yet
- Notes (AH-302-ABG-Mid)Document12 pagesNotes (AH-302-ABG-Mid)Hamza Bin SajidNo ratings yet
- Week 3 GeneticsDocument16 pagesWeek 3 GeneticsRich Darlene Dela CruzNo ratings yet
- A. Origin Importance of GeneticsDocument37 pagesA. Origin Importance of GeneticsArmie Joy ManingatNo ratings yet
- Fish Genetics & BreedingDocument237 pagesFish Genetics & BreedingPurva SharanNo ratings yet
- 8 GeneticsDocument85 pages8 GeneticsRoselyn CarmenNo ratings yet
- Nucleotides, nucleic acids and the genetic material explainedDocument41 pagesNucleotides, nucleic acids and the genetic material explainedMario Mallare AbesamisNo ratings yet
- Heredity and Evolution Class 10 Notes CBSE Science Chapter 9 PDFDocument11 pagesHeredity and Evolution Class 10 Notes CBSE Science Chapter 9 PDFvanshNo ratings yet
- Class - X - Science - Heredity and Evolution - PPT PDFDocument11 pagesClass - X - Science - Heredity and Evolution - PPT PDFAvrian PradiptyaNo ratings yet
- 1.GM ClassDocument74 pages1.GM ClassChandrachur GhoshNo ratings yet
- 2 18KP2B06 2021012801523485Document45 pages2 18KP2B06 2021012801523485Rey OrbeNo ratings yet
- CBSE Class 12 Biology Chapter 5 Principles of Inheritance and Variation Revision NotesDocument35 pagesCBSE Class 12 Biology Chapter 5 Principles of Inheritance and Variation Revision NotesAnkur YadavNo ratings yet
- MODULE 1-GeneticsDocument13 pagesMODULE 1-GeneticsDivine Grace CincoNo ratings yet
- Life Begins With A CellDocument23 pagesLife Begins With A Cellva1612315No ratings yet
- 3- An IntromoltocellDocument48 pages3- An Intromoltocellbibi.hansmuddyNo ratings yet
- Genes Environment AND DevelopmentDocument29 pagesGenes Environment AND Developmentsushmita bhartiaNo ratings yet
- Classical (Mendelian) GeneticsDocument32 pagesClassical (Mendelian) GeneticsKOMAL FATIMANo ratings yet
- GeneticsDocument33 pagesGeneticsPrashna KoiralaNo ratings yet
- Biol 12 Genetics Notes and University Entrance Exam Questions 2003Document66 pagesBiol 12 Genetics Notes and University Entrance Exam Questions 2003Tizita EmanaNo ratings yet
- Mendelian Genetic DisordersDocument8 pagesMendelian Genetic DisordersPrajwal Dixit0% (1)
- Mendel's Pea Plant Experiments Reveal Inheritance PatternsDocument12 pagesMendel's Pea Plant Experiments Reveal Inheritance PatternsMaryfleur BazarNo ratings yet
- Unit 7 HeredityDocument20 pagesUnit 7 HeredityAmriani AnrahNo ratings yet
- Chapter 1.1Document70 pagesChapter 1.1AS2012B2 FATIN NUR AQILAH BINTI DAZALINo ratings yet
- Bio 97 Genetics Lecture NotesDocument5 pagesBio 97 Genetics Lecture Notesstevom188No ratings yet
- Notes On CBSE 5Document88 pagesNotes On CBSE 5Marvin ArcillaNo ratings yet
- Cytogenetics Compiled Topic Notes Topic 1: Origin and Importance of CytogeneticsDocument10 pagesCytogenetics Compiled Topic Notes Topic 1: Origin and Importance of CytogeneticscassseeeyyyNo ratings yet
- KumawatDocument67 pagesKumawatpiyushjakhar05No ratings yet
- Introduction To Genetics Study GuideDocument67 pagesIntroduction To Genetics Study GuideDanica Lao-ayNo ratings yet
- Biology of Life and GeneticsDocument59 pagesBiology of Life and Geneticsjawahar100% (1)
- The Beginnings of Genetics - REPORT BY STUDENTS OF 10ADocument18 pagesThe Beginnings of Genetics - REPORT BY STUDENTS OF 10AFawwazNo ratings yet
- GeneticsDocument25 pagesGeneticsLouel Mattew LagascaNo ratings yet
- Mendelian Genetic DisordersDocument8 pagesMendelian Genetic DisordersGobinda Munda100% (1)
- Lec5 Investigation Approach To Genetics IDocument24 pagesLec5 Investigation Approach To Genetics ISITI BAZILAH BINTI BILAK KPM-GuruNo ratings yet
- 1history of Genetics 1Document8 pages1history of Genetics 1Catherine Noynay RaagasNo ratings yet
- Pre Post MendelianDocument5 pagesPre Post Mendeliansakshi sharmaNo ratings yet
- Unit 3 Bio4Er NotesDocument42 pagesUnit 3 Bio4Er NoteskgpunithaNo ratings yet
- Genetic Linkage Is A Term Which Describes The Tendency of Certain Loci or Alleles To BeDocument34 pagesGenetic Linkage Is A Term Which Describes The Tendency of Certain Loci or Alleles To BeHabibur RahamanNo ratings yet
- MendelDocument15 pagesMendelAfaq AhmadNo ratings yet
- ANTONIO - Module 1 AssessmentDocument2 pagesANTONIO - Module 1 AssessmentJazMine AntonioNo ratings yet
- Principles of Inheritance and Variation (Genetics - I)Document34 pagesPrinciples of Inheritance and Variation (Genetics - I)malavikaNo ratings yet
- Introduction To GeneticsDocument38 pagesIntroduction To GeneticsCaptain WolfNo ratings yet
- Hybrids: Mendel's GeneticsDocument5 pagesHybrids: Mendel's GeneticsAnonymous doCtd0IJDNNo ratings yet
- DNA as the Genetic MaterialDocument111 pagesDNA as the Genetic MaterialSrinath ShankarNo ratings yet
- Insect Biochemistry and Molecular Biology: Giuseppe SacconeDocument32 pagesInsect Biochemistry and Molecular Biology: Giuseppe SacconeTrya CahyantiNo ratings yet
- CBSE Class 10 Science Notes Chapter 9 Heredity and EvolutionDocument18 pagesCBSE Class 10 Science Notes Chapter 9 Heredity and EvolutionHarsh AgarwalNo ratings yet
- Genetics 1.1 ResumeDocument102 pagesGenetics 1.1 ResumemiertschinkNo ratings yet
- Genetics in Orthodontics - 12 11 23Document88 pagesGenetics in Orthodontics - 12 11 23dr.sakshigarg0105No ratings yet
- MedilianismDocument16 pagesMedilianismYujin AhnNo ratings yet
- 9 - Notes HEREDITYDocument5 pages9 - Notes HEREDITYSuraj BhandareNo ratings yet
- Lecture Genetics Science of Heredity and VariationDocument31 pagesLecture Genetics Science of Heredity and VariationGideon CavidaNo ratings yet
- Principles of Genetics: Module - 3Document26 pagesPrinciples of Genetics: Module - 3Indrojyoti MondalNo ratings yet
- Lesson 6Document40 pagesLesson 6charith chiranthaNo ratings yet
- Lesson 5Document63 pagesLesson 5charith chiranthaNo ratings yet
- Lesson 4 Transcription in Eukaryotic CellsDocument66 pagesLesson 4 Transcription in Eukaryotic Cellscharith chiranthaNo ratings yet
- Lesson 3: Transcription in BacteriaDocument63 pagesLesson 3: Transcription in Bacteriacharith chiranthaNo ratings yet
- Lesson 2Document62 pagesLesson 2charith chiranthaNo ratings yet
- Chanel SWOT AnalysisDocument5 pagesChanel SWOT AnalysisJeish KimNo ratings yet
- Admission Procedure For International StudentsDocument8 pagesAdmission Procedure For International StudentsAndreea Anghel-DissanayakaNo ratings yet
- 99th Indian Science Congress (Bhubaneshwar)Document95 pages99th Indian Science Congress (Bhubaneshwar)Aadarsh DasNo ratings yet
- History and Development of the Foodservice IndustryDocument23 pagesHistory and Development of the Foodservice IndustryMaria Athenna MallariNo ratings yet
- Phaseo Abl7 Abl8 Abl8rps24100Document9 pagesPhaseo Abl7 Abl8 Abl8rps24100Magda DiazNo ratings yet
- Final - WPS PQR 86Document4 pagesFinal - WPS PQR 86Parag WadekarNo ratings yet
- PediculosisDocument14 pagesPediculosisREYMARK HACOSTA100% (1)
- PBS-P100 Facilities Standards GuideDocument327 pagesPBS-P100 Facilities Standards Guidecessna5538cNo ratings yet
- Appendix 1 Application FormDocument13 pagesAppendix 1 Application FormSharifahrodiah SemaunNo ratings yet
- 20ME901 Automobile Engineering Unit 3Document74 pages20ME901 Automobile Engineering Unit 36044 sriramNo ratings yet
- Geometric Sculpture From 72 PencilsDocument5 pagesGeometric Sculpture From 72 PencilsHugo Iván Gonzalez CruzNo ratings yet
- Presentation 1Document20 pagesPresentation 1nikitakhanduja1304No ratings yet
- Stages of Intimate RelationshipsDocument4 pagesStages of Intimate RelationshipsKrystalline ParkNo ratings yet
- Analysis and Design of Multi Storey Building by Using STAAD ProDocument5 pagesAnalysis and Design of Multi Storey Building by Using STAAD Prolikith rockNo ratings yet
- Oracle® E-Business Suite: Integrated SOA Gateway Implementation Guide Release 12.2Document202 pagesOracle® E-Business Suite: Integrated SOA Gateway Implementation Guide Release 12.2yadavdevenderNo ratings yet
- Soal Kelas 4 IIDocument5 pagesSoal Kelas 4 IIes tougeNo ratings yet
- PiXL Knowledge Test ANSWERS - AQA B1 CORE Science - Legacy (2016 and 2017)Document12 pagesPiXL Knowledge Test ANSWERS - AQA B1 CORE Science - Legacy (2016 and 2017)Mrs S BakerNo ratings yet
- Fjords Blue Web ENG PDFDocument20 pagesFjords Blue Web ENG PDFMoldovan MirceaNo ratings yet
- Class 7thDocument24 pagesClass 7thPriyaNo ratings yet
- Serendipity - A Sociological NoteDocument2 pagesSerendipity - A Sociological NoteAmlan BaruahNo ratings yet
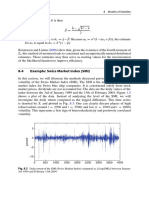
- 8.4 Example: Swiss Market Index (SMI) : 188 8 Models of VolatilityDocument3 pages8.4 Example: Swiss Market Index (SMI) : 188 8 Models of VolatilityNickesh ShahNo ratings yet
- ProjectDocument86 pagesProjectrajuNo ratings yet
- C11 Strategy DevelopmentDocument30 pagesC11 Strategy DevelopmentPARTI KEADILAN RAKYAT NIBONG TEBALNo ratings yet
- Best Home Oxygen Concentrators-Lowest Prices & Fast Shipping (Oxygen Machines) - 2021 - YuwellDocument1 pageBest Home Oxygen Concentrators-Lowest Prices & Fast Shipping (Oxygen Machines) - 2021 - YuwellPelayanan ResusitasiNo ratings yet
- Year 10 Maths PlaneDocument62 pagesYear 10 Maths Planehal wangNo ratings yet
- Agilent Cool On-Column Operation ManualDocument42 pagesAgilent Cool On-Column Operation Manualdmcevoy1965No ratings yet
- Medicinal PlantDocument13 pagesMedicinal PlantNeelum iqbalNo ratings yet
- Chirag STDocument18 pagesChirag STchiragNo ratings yet
- Chronological OrderDocument5 pagesChronological OrderDharWin d'Wing-Wing d'AriestBoyzNo ratings yet