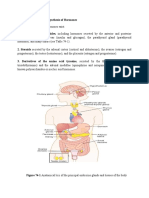
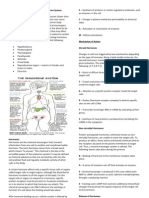
You might also like
- Prof. N. Syabbalo: MB., CHB., PHD., FCCP., FRS., Fiba Professor of Physiology & MedicineDocument45 pagesProf. N. Syabbalo: MB., CHB., PHD., FCCP., FRS., Fiba Professor of Physiology & MedicineHomeground entertainmentNo ratings yet
- Hormone: For Other Uses, SeeDocument5 pagesHormone: For Other Uses, SeeEki Ryan SetiowatiNo ratings yet
- Adrenocorticotropic Hormone (ACTH) From The Anterior Pituitary Gland Specifically StimulatesDocument2 pagesAdrenocorticotropic Hormone (ACTH) From The Anterior Pituitary Gland Specifically StimulatesStjepan ErženNo ratings yet
- Unit 1 Courselink NotesDocument27 pagesUnit 1 Courselink NotesLauren StamNo ratings yet
- Hormonal ActionDocument37 pagesHormonal ActionHomeground entertainmentNo ratings yet
- Study of The Endocrine SystemDocument225 pagesStudy of The Endocrine SystemHuzaifa RashidNo ratings yet
- Endrocrinology - PPTX Version 1Document40 pagesEndrocrinology - PPTX Version 1Abishek BhadraNo ratings yet
- Hormone: For Other Uses, SeeDocument4 pagesHormone: For Other Uses, SeePrasanna Nair ParameswaranNo ratings yet
- What Are HormonesDocument16 pagesWhat Are HormonesHarry RoyNo ratings yet
- The Endocrine System: A System of Chemical CommunicationDocument7 pagesThe Endocrine System: A System of Chemical CommunicationKaleem UllahNo ratings yet
- Biochemistry of HormonDocument16 pagesBiochemistry of HormonMuhamad SdeqNo ratings yet
- Hormones, ClassificationDocument10 pagesHormones, ClassificationMenoNo ratings yet
- EndocrinologyDocument35 pagesEndocrinologyNasif HussainNo ratings yet
- 1 - Introduction EndocrinologyDocument40 pages1 - Introduction EndocrinologyBHUWAN BASKOTANo ratings yet
- The Endocrine System: Hormones, Receptors and RegulationDocument47 pagesThe Endocrine System: Hormones, Receptors and RegulationRifi OktasiaNo ratings yet
- Chemistry ProjectDocument23 pagesChemistry Projecttwinklelee123450% (2)
- Chemical Structure and Synthesis of HormonesDocument6 pagesChemical Structure and Synthesis of HormonesFAHEEM UD DINNo ratings yet
- Endocrine System AssesmentDocument51 pagesEndocrine System AssesmentYudiatma100% (1)
- Endocrine Physiology Lecture 2Document9 pagesEndocrine Physiology Lecture 2Tofunmi Kunle-KunbiNo ratings yet
- Introduction To Endocrinology: Dr. Jehad Al-ShuneigatDocument21 pagesIntroduction To Endocrinology: Dr. Jehad Al-Shuneigatyousef sarairehNo ratings yet
- 4 Endocrine 1Document18 pages4 Endocrine 1ShenNo ratings yet
- Endocrine Anatomy and PhysiologyDocument10 pagesEndocrine Anatomy and PhysiologyJojo JustoNo ratings yet
- Biology ProjectDocument27 pagesBiology Projectravifact0% (2)
- SGD Physiology Endocrine and MetabolismDocument7 pagesSGD Physiology Endocrine and MetabolismTinesh RajahNo ratings yet
- ABS Endocrinology PDFDocument59 pagesABS Endocrinology PDFMichael Olivier WNo ratings yet
- EndocrinologyDocument223 pagesEndocrinologyMr mk RollinsNo ratings yet
- SUMAN - TAMANG2021-02-14Regulation of Hormone ActionDocument43 pagesSUMAN - TAMANG2021-02-14Regulation of Hormone Actions vNo ratings yet
- Coordination of Body Functions by Chemical MessengerDocument6 pagesCoordination of Body Functions by Chemical MessengerHorain FatimaNo ratings yet
- Introduction - CC3: Mechanism of Hormonal ActionDocument40 pagesIntroduction - CC3: Mechanism of Hormonal ActionAl-hadad AndromacheNo ratings yet
- Endocrinology 1st TrimesterDocument16 pagesEndocrinology 1st TrimesteriwennieNo ratings yet
- Hormonal Management of Male InfertilityDocument22 pagesHormonal Management of Male Infertilitydranibalurologoencancun.mxNo ratings yet
- Chapter - 17 PhysiologyDocument49 pagesChapter - 17 PhysiologyAhsanullah PathanNo ratings yet
- Endocrine SystemDocument34 pagesEndocrine SystemNada DardeerNo ratings yet
- Hormones:: Signaling MoleculesDocument20 pagesHormones:: Signaling MoleculesSangeeta DwivediNo ratings yet
- Endocrine SystemDocument41 pagesEndocrine SystemMerrin EncarnacionNo ratings yet
- Endocrine System OutlineDocument17 pagesEndocrine System OutlineEsthermae PanadenNo ratings yet
- E N D o C R I N o L o G Y: Dr. Victoria G. Giango Chair, Dept. of PhysiologyDocument23 pagesE N D o C R I N o L o G Y: Dr. Victoria G. Giango Chair, Dept. of Physiologyanon-271548No ratings yet
- Lec 8 - Hormones PDFDocument13 pagesLec 8 - Hormones PDFrajeshNo ratings yet
- Hormones EssayDocument2 pagesHormones Essayapi-254545870No ratings yet
- Learning JournalDocument82 pagesLearning JournalAnnapurna RavelNo ratings yet
- Anatomy and Physiology-Endocrine SystemDocument5 pagesAnatomy and Physiology-Endocrine SystemEixid Enna YeLikNo ratings yet
- Overview of Endocrine SystemDocument16 pagesOverview of Endocrine System2023ph17No ratings yet
- HORMONESDocument9 pagesHORMONESSARA M SNo ratings yet
- UntitledDocument12 pagesUntitledDerrick kinyaNo ratings yet
- HormonesDocument10 pagesHormonesAmjad AlmousawiNo ratings yet
- Endocrine System CommunicationDocument32 pagesEndocrine System CommunicationalallaallalaNo ratings yet
- Transport of HormonesDocument5 pagesTransport of HormonesJohn MusaNo ratings yet
- 6 Group Phsiology.....Document14 pages6 Group Phsiology.....Ayat ChaudaryNo ratings yet
- Care of Clients With Problem On EndocrineDocument48 pagesCare of Clients With Problem On EndocrineAngel CauilanNo ratings yet
- Biochemistry Assignment Submitted To: Dr. Noureen Submitted byDocument15 pagesBiochemistry Assignment Submitted To: Dr. Noureen Submitted bySalman KhanNo ratings yet
- Endocrinology PDFDocument11 pagesEndocrinology PDFLawrence Genelago GamboaNo ratings yet
- Introduction To Endocrine Physiology: DR EvaDocument17 pagesIntroduction To Endocrine Physiology: DR EvaEmmanuel Julius ChalighaNo ratings yet
- Endocrine PhysiologyDocument41 pagesEndocrine PhysiologyRaj CellaNo ratings yet
- Key Concepts of EndocrinologyDocument131 pagesKey Concepts of Endocrinologykomal pattabiNo ratings yet
- The Endocrine System: Hormone Classification and Mechanisms of Intercellular CommunicationDocument82 pagesThe Endocrine System: Hormone Classification and Mechanisms of Intercellular Communicationعم رNo ratings yet
- Endocrine SystemDocument22 pagesEndocrine Systempranutan739No ratings yet
- Lecture 2 Endocrine SystemDocument53 pagesLecture 2 Endocrine SystemLouella ArtatesNo ratings yet
- ENDOCRINE SYSTEM - Anatomy and PhysiologyDocument10 pagesENDOCRINE SYSTEM - Anatomy and PhysiologyJay Crishnan Morales CajandingNo ratings yet
- Endocrine System: A Tutorial Study GuideFrom EverandEndocrine System: A Tutorial Study GuideRating: 5 out of 5 stars5/5 (1)
- Cesarean SectionDocument36 pagesCesarean SectionHomeground entertainmentNo ratings yet
- Breech PresentationDocument27 pagesBreech PresentationHomeground entertainmentNo ratings yet
- 12558931Document34 pages12558931Homeground entertainmentNo ratings yet
- Cervical CancerDocument87 pagesCervical CancerHomeground entertainmentNo ratings yet
- CaesareanDocument32 pagesCaesareanHomeground entertainmentNo ratings yet
- Perinatal Asphyxia GuideDocument37 pagesPerinatal Asphyxia GuideHomeground entertainmentNo ratings yet
- Amino Acids: Eric Mbindo NjunjuDocument18 pagesAmino Acids: Eric Mbindo NjunjuHomeground entertainmentNo ratings yet
- Biochemistry MBS 230: Eric Mbindo NjunjuDocument20 pagesBiochemistry MBS 230: Eric Mbindo NjunjuHomeground entertainmentNo ratings yet
- Convulsive Disorders: Presenters: Kabwe Chanda EliasDocument30 pagesConvulsive Disorders: Presenters: Kabwe Chanda EliasHomeground entertainmentNo ratings yet
- Drug Treatment of Arrhythmias-Feb 2015Document39 pagesDrug Treatment of Arrhythmias-Feb 2015Homeground entertainmentNo ratings yet
- Proteins: Eric Mbindo NjunjuDocument76 pagesProteins: Eric Mbindo NjunjuHomeground entertainmentNo ratings yet
- Mutations and Their OriginsDocument62 pagesMutations and Their OriginsHomeground entertainmentNo ratings yet
- P6 Disorders of The Basal GangliaDocument12 pagesP6 Disorders of The Basal GangliaHomeground entertainmentNo ratings yet
- Biochemistry MBS 230: Eric Mbindo NjunjuDocument20 pagesBiochemistry MBS 230: Eric Mbindo NjunjuHomeground entertainmentNo ratings yet
- DRUG TREATMENT OF BRONCHIAL ASTHMADocument39 pagesDRUG TREATMENT OF BRONCHIAL ASTHMAHomeground entertainmentNo ratings yet
- Anaphylaxis: Copperbelt University School of Medicine MBCHB Pharmacology February 2015 (DR Sindwa Kanyimba)Document12 pagesAnaphylaxis: Copperbelt University School of Medicine MBCHB Pharmacology February 2015 (DR Sindwa Kanyimba)Homeground entertainmentNo ratings yet
- Principles of Diet Diversity and Dietary Modification - 2Document46 pagesPrinciples of Diet Diversity and Dietary Modification - 2Homeground entertainmentNo ratings yet
- p4 Extra Pyramidal SystemDocument19 pagesp4 Extra Pyramidal SystemHomeground entertainmentNo ratings yet
- p8 The Physiology of The Flocculonodular LobeDocument16 pagesp8 The Physiology of The Flocculonodular LobeHomeground entertainmentNo ratings yet
- p5 Basal GangliaDocument18 pagesp5 Basal GangliaHomeground entertainmentNo ratings yet
- Anti-Tussives-Mucolytics-Expectorants-Feb 2015Document10 pagesAnti-Tussives-Mucolytics-Expectorants-Feb 2015Homeground entertainmentNo ratings yet
- p5 Basal GangliaDocument18 pagesp5 Basal GangliaHomeground entertainmentNo ratings yet
- P9 Disorders of The CerebellumDocument29 pagesP9 Disorders of The CerebellumHomeground entertainmentNo ratings yet
- P3 Pyramidal Tracts of The Somatomotor SystemDocument17 pagesP3 Pyramidal Tracts of The Somatomotor SystemHomeground entertainmentNo ratings yet
- p8 The Physiology of The Flocculonodular LobeDocument16 pagesp8 The Physiology of The Flocculonodular LobeHomeground entertainmentNo ratings yet
- P9 Disorders of The CerebellumDocument29 pagesP9 Disorders of The CerebellumHomeground entertainmentNo ratings yet
- P2 Cortical Areas and Their Role in Contral of MovementDocument18 pagesP2 Cortical Areas and Their Role in Contral of MovementHomeground entertainmentNo ratings yet
- P6 Disorders of The Basal GangliaDocument12 pagesP6 Disorders of The Basal GangliaHomeground entertainmentNo ratings yet
- p4 Extra Pyramidal SystemDocument19 pagesp4 Extra Pyramidal SystemHomeground entertainmentNo ratings yet
- Neurophysiology Systems of the Brain and Spinal CordDocument50 pagesNeurophysiology Systems of the Brain and Spinal CordHomeground entertainmentNo ratings yet
- What Is Mean?: Extrapolation InterpolationDocument2 pagesWhat Is Mean?: Extrapolation InterpolationVinod SharmaNo ratings yet
- Foundations On Expansive Soils: 3.1. BackgroundDocument31 pagesFoundations On Expansive Soils: 3.1. BackgroundbiniNo ratings yet
- Mark Dyczkowski and Trika Journal March 2015 Vol.1.No.1.Document10 pagesMark Dyczkowski and Trika Journal March 2015 Vol.1.No.1.Mark Dyczkoswki and Trika Journal100% (2)
- But Virgil Was Not There": The Lasting Impact of Dante's Homosocial HellDocument7 pagesBut Virgil Was Not There": The Lasting Impact of Dante's Homosocial HellЮлия ЧебанNo ratings yet
- DANGEL 4as LESSON PLANNINGDocument2 pagesDANGEL 4as LESSON PLANNINGCarlz BrianNo ratings yet
- Neolithic Farming Villages Jericho and Catal HuyukDocument1 pageNeolithic Farming Villages Jericho and Catal HuyukPream BoleoNo ratings yet
- Battle Bikes 2.4 PDFDocument56 pagesBattle Bikes 2.4 PDFfranzyland100% (1)
- IM PS Fashion-Business-Digital-Communication-And-Media 3Y Course Pathway MI 04Document7 pagesIM PS Fashion-Business-Digital-Communication-And-Media 3Y Course Pathway MI 04oliwia bujalskaNo ratings yet
- Ffective Riting Kills: Training & Discussion OnDocument37 pagesFfective Riting Kills: Training & Discussion OnKasi ReddyNo ratings yet
- 2011 Mena Annual Reportv1Document73 pages2011 Mena Annual Reportv1Yasmeen LayallieNo ratings yet
- 17a03g - Mosfet - DualDocument5 pages17a03g - Mosfet - DualEletronica01 - BLUEVIXNo ratings yet
- Vocabulary Practice 1Document3 pagesVocabulary Practice 1Phuong AnhNo ratings yet
- The Neuroscience of Autism Spectrum DisordersDocument10 pagesThe Neuroscience of Autism Spectrum DisorderssouciNo ratings yet
- James M. Buchanan - Why I, Too, Am Not A Conservative PDFDocument121 pagesJames M. Buchanan - Why I, Too, Am Not A Conservative PDFHeitor Berbigier Bandas100% (2)
- 1161 Article+Text 5225 1 4 20220712Document9 pages1161 Article+Text 5225 1 4 20220712Warman FatraNo ratings yet
- Samruddhi ComplexDocument7 pagesSamruddhi ComplexNews Side Effects.No ratings yet
- Robotics Process AutomationDocument21 pagesRobotics Process Automationbhaskarkiran.pNo ratings yet
- Mercer Role and Job Analysis InfoDocument3 pagesMercer Role and Job Analysis InfojehaniaNo ratings yet
- MsdsDocument6 pagesMsdsGis GeorgeNo ratings yet
- PW 160-Taliban Fragmentation Fact Fiction and Future-PwDocument28 pagesPW 160-Taliban Fragmentation Fact Fiction and Future-Pwrickyricardo1922No ratings yet
- Basic Load (Individual) Veterinarian Field PackDocument3 pagesBasic Load (Individual) Veterinarian Field PackJohn MillerNo ratings yet
- Ceftriaxone R Salmonella Typhi 02Document8 pagesCeftriaxone R Salmonella Typhi 02docsumitraiNo ratings yet
- ErgonomicsDocument15 pagesErgonomicsdtmNo ratings yet
- 2020 Exam-Sample-Questions-Computer-ScienceDocument8 pages2020 Exam-Sample-Questions-Computer-ScienceNesrine LaradjiNo ratings yet
- Combining Singing and PsycologyDocument6 pagesCombining Singing and PsycologyAna luciaNo ratings yet
- Mikes ResumeDocument2 pagesMikes Resumeapi-312645878No ratings yet
- Four Pillars of EducationDocument42 pagesFour Pillars of EducationWinter BacalsoNo ratings yet
- CBSE Class 10 Science Revision Notes Chapter - 2 Acids, Bases and SaltsDocument11 pagesCBSE Class 10 Science Revision Notes Chapter - 2 Acids, Bases and Saltsmilind dhamaniyaNo ratings yet
- New Balance Case StudyDocument3 pagesNew Balance Case StudyDimas AdityaNo ratings yet
- 5G Antenna Talk TWDocument48 pages5G Antenna Talk TWRohit MathurNo ratings yet