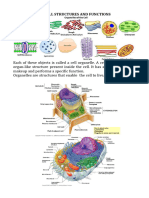
You might also like
- Principles, Applications and InterpretationsDocument64 pagesPrinciples, Applications and InterpretationsAbbi Yanto ArtNo ratings yet
- Innovative Solutions For Flow Cytometry Analysis Optimized Kits and Reagents PB3322ENEUDocument32 pagesInnovative Solutions For Flow Cytometry Analysis Optimized Kits and Reagents PB3322ENEUoscar_rogeroNo ratings yet
- Bacterial Cell Structure: By: Hafiza Asfa Shafique Microbiology BS Biotechnology VDocument30 pagesBacterial Cell Structure: By: Hafiza Asfa Shafique Microbiology BS Biotechnology VJawad100% (3)
- Learning Activity Sheet Biotechnology 8Document11 pagesLearning Activity Sheet Biotechnology 8Jaeda BaltazarNo ratings yet
- Cambridge Science Worksheet 8Document13 pagesCambridge Science Worksheet 8ozmanNo ratings yet
- General Biology 1 QuizDocument2 pagesGeneral Biology 1 QuizQueng ElediaNo ratings yet
- Molecular Cell Biology Lodish 7th Edition Solutions ManualDocument8 pagesMolecular Cell Biology Lodish 7th Edition Solutions ManualVictoriaWilliamsegtnm100% (83)
- Synthetic Biological Circuit Design Implementing Protein Degradation In-VitroDocument5 pagesSynthetic Biological Circuit Design Implementing Protein Degradation In-VitroRohit SharmaNo ratings yet
- Sampathkumar Proteins-StructFunctBioinform 2011Document6 pagesSampathkumar Proteins-StructFunctBioinform 2011ctak2022No ratings yet
- Parte 03Document100 pagesParte 03Pâmella PicançoNo ratings yet
- Lovely Professional University: Assignment ReportDocument5 pagesLovely Professional University: Assignment ReportAdarshNo ratings yet
- TMP 324 BDocument17 pagesTMP 324 BnithiananthiNo ratings yet
- Chapter 3 Subcellular Fractionation and Isolation of OrganellesDocument8 pagesChapter 3 Subcellular Fractionation and Isolation of OrganellesrajaNo ratings yet
- Induction of Apoptotic Program in Cell-Free Extracts: Requirement For dATP and Cytochrome CDocument11 pagesInduction of Apoptotic Program in Cell-Free Extracts: Requirement For dATP and Cytochrome CJaja LazaroNo ratings yet
- Jove 112 54024Document10 pagesJove 112 54024Nadim AlbirNo ratings yet
- Nucleus 2015Document24 pagesNucleus 2015Analiz De Jesús LópezNo ratings yet
- Cell Biology: Course Code: LSE-01 Assignment Code: LSE-01/TMA/2020 Maximum Marks: 100Document26 pagesCell Biology: Course Code: LSE-01 Assignment Code: LSE-01/TMA/2020 Maximum Marks: 100Rajni KumariNo ratings yet
- Monocle VignetteDocument13 pagesMonocle VignetteToh Qin KaneNo ratings yet
- Dissertation Talk7Document39 pagesDissertation Talk7Chris McGeeNo ratings yet
- Possible Mechanism of Porosome-Mediated Exosome Release: Key Terms: Neuronal Porosome, Beta CellDocument30 pagesPossible Mechanism of Porosome-Mediated Exosome Release: Key Terms: Neuronal Porosome, Beta CellValen EstevezNo ratings yet
- Name REEMA AMIN Registration No FA19Document5 pagesName REEMA AMIN Registration No FA19reema amynNo ratings yet
- 9 Flow Cytometric Strategies To Study CNS DevelopmentDocument32 pages9 Flow Cytometric Strategies To Study CNS DevelopmenttaroNo ratings yet
- Analysis of Monoclonal Immunoglobulins From Bone Marrow Plasma CellsDocument9 pagesAnalysis of Monoclonal Immunoglobulins From Bone Marrow Plasma CellsSabrina DaidNo ratings yet
- tmp992F TMPDocument10 pagestmp992F TMPFrontiersNo ratings yet
- TSE Method of Periplasmic Protein ExtractionDocument8 pagesTSE Method of Periplasmic Protein ExtractionAditya BasuNo ratings yet
- 2 BM 2008-09Document26 pages2 BM 2008-09Nagham Bazzi100% (2)
- Annual Reviews - ReplicationDocument36 pagesAnnual Reviews - ReplicationfngermanoNo ratings yet
- Protein Chips and Functional ProteomicsDocument9 pagesProtein Chips and Functional ProteomicsAmal ..No ratings yet
- Worksheet 2 The Cell WalaDocument7 pagesWorksheet 2 The Cell WalacalliemozartNo ratings yet
- Aggregating Brain Cell Cultures For Neurotoxicological StudiesDocument24 pagesAggregating Brain Cell Cultures For Neurotoxicological Studiesmarej312No ratings yet
- Nucleation of Sub-Micrometer Protein Crystals in Square-Shaped Macroporous Silicon StructuresDocument8 pagesNucleation of Sub-Micrometer Protein Crystals in Square-Shaped Macroporous Silicon StructuresNoé Luna JimNo ratings yet
- JCB 202209062Document21 pagesJCB 202209062Aleksay NesmashnyyNo ratings yet
- Summers 2023Document19 pagesSummers 2023Pilar AufrastoNo ratings yet
- European Journal of Biochemistry - 2001 - Kalies - Protein Translocation Into The Endoplasmic Reticulum ERDocument5 pagesEuropean Journal of Biochemistry - 2001 - Kalies - Protein Translocation Into The Endoplasmic Reticulum ER12 yashika palNo ratings yet
- Human Plasma PeptideAtlasDocument4 pagesHuman Plasma PeptideAtlasJheny LopezNo ratings yet
- Single Cell Mass SpectrometryDocument4 pagesSingle Cell Mass SpectrometryDave ZNo ratings yet
- Protein Microarray - WikipediaDocument10 pagesProtein Microarray - Wikipediasakib sadmanNo ratings yet
- Molecular Biology of The Cell 5th Edition Alberts Test BankDocument13 pagesMolecular Biology of The Cell 5th Edition Alberts Test Bankodettedieupmx23m100% (31)
- Neurochemistry International: Oliver Maier, Julia Böhm, Michael Dahm, Stefan Brück, Cordian Beyer, Sonja JohannDocument10 pagesNeurochemistry International: Oliver Maier, Julia Böhm, Michael Dahm, Stefan Brück, Cordian Beyer, Sonja JohannPili CárdenasNo ratings yet
- Methods of Protein AnalysisDocument41 pagesMethods of Protein AnalysisObin MrazNo ratings yet
- Organic Sensors 1Document158 pagesOrganic Sensors 1るよNo ratings yet
- 1600830Document18 pages1600830teliatchagou12No ratings yet
- Molecular Biology of The Cell 5Th Edition Alberts Test Bank Full Chapter PDFDocument34 pagesMolecular Biology of The Cell 5Th Edition Alberts Test Bank Full Chapter PDFrorybridgetewe100% (8)
- Isolation of Kuppfer CellsDocument9 pagesIsolation of Kuppfer CellsusmanNo ratings yet
- In Silico Attributes of Cold Resistance Protein 1 (Crp1) F Rom Brassica OleraceaeDocument8 pagesIn Silico Attributes of Cold Resistance Protein 1 (Crp1) F Rom Brassica OleraceaeresearchinventyNo ratings yet
- Cell Sorting ThesisDocument5 pagesCell Sorting Thesissusanmatthewssouthbend100% (2)
- Zyuzin Et Al. - 2022 - Biodegradable Particles For Protein Delivery EstiDocument9 pagesZyuzin Et Al. - 2022 - Biodegradable Particles For Protein Delivery Estiquentin pNo ratings yet
- Electrophysiological Recording Techniques PDFDocument7 pagesElectrophysiological Recording Techniques PDFAndrei TatomirNo ratings yet
- Apoptosis PlaybookDocument20 pagesApoptosis PlaybookArmando SignoreNo ratings yet
- 2011 TA Insertion Into ERDocument12 pages2011 TA Insertion Into ERLê Khánh ToànNo ratings yet
- Chapter 06Document39 pagesChapter 06NnleinomNo ratings yet
- Prinsip Sysmex Hematology AnalyzerDocument5 pagesPrinsip Sysmex Hematology AnalyzerLince Wijoyo100% (1)
- Attachment-54605872 6Document5 pagesAttachment-54605872 6Vania Nurul SabarinaNo ratings yet
- A Few Approaches To The Study of Endomembranes: Insights Gained From AutoradiographyDocument10 pagesA Few Approaches To The Study of Endomembranes: Insights Gained From AutoradiographyDanie MorenoNo ratings yet
- A Cell Biology Laboratory Exercise To Study Sub-Cellular OrganellDocument6 pagesA Cell Biology Laboratory Exercise To Study Sub-Cellular OrganellMarianita MontoyaNo ratings yet
- 824-Texto Del Manuscrito Completo (Cuadros y Figuras Insertos) - 4381-1!10!20120828Document6 pages824-Texto Del Manuscrito Completo (Cuadros y Figuras Insertos) - 4381-1!10!20120828Tallie ZeidlerNo ratings yet
- Golgi Outposts and Satellites in Neurons - Secretory Trafficking in Neuronal DendritesDocument7 pagesGolgi Outposts and Satellites in Neurons - Secretory Trafficking in Neuronal DendritesEshaNo ratings yet
- Integrative CUT&Tag-RNA-Seq Analysis of Histone Variant macroH2A1-dependent Orchestration of Human Induced Pluripotent Stem Cell ReprogrammingDocument16 pagesIntegrative CUT&Tag-RNA-Seq Analysis of Histone Variant macroH2A1-dependent Orchestration of Human Induced Pluripotent Stem Cell Reprogrammingniccolo.liorni1No ratings yet
- Molecular Cell Biology 7th Edition Lodish Test BankDocument8 pagesMolecular Cell Biology 7th Edition Lodish Test Bankrosaanqo8100% (33)
- Brouzes 2009Document6 pagesBrouzes 2009Salman AhmadNo ratings yet
- Protein Engineering Reference ArticleDocument27 pagesProtein Engineering Reference ArticleSiddhant BanwatNo ratings yet
- The Cell: Anatomy and Division: ExerciseDocument4 pagesThe Cell: Anatomy and Division: ExerciseMarielle SaymanNo ratings yet
- Kinetochore Assembly and Function Through The Cell Cycle: ReviewDocument15 pagesKinetochore Assembly and Function Through The Cell Cycle: ReviewLê Khánh ToànNo ratings yet
- Virology Journal: Predicting The Subcellular Localization of Viral Proteins Within A Mammalian Host CellDocument8 pagesVirology Journal: Predicting The Subcellular Localization of Viral Proteins Within A Mammalian Host CellRuy Lopez ClosedNo ratings yet
- Gram-Positive and Gram-Negative Subcellular Localization Using Rotation Forest and Physicochemical-Based FeaturesDocument8 pagesGram-Positive and Gram-Negative Subcellular Localization Using Rotation Forest and Physicochemical-Based FeaturesRuy Lopez ClosedNo ratings yet
- Standard Tries - Compressed Tries - Suffix TriesDocument11 pagesStandard Tries - Compressed Tries - Suffix TriesRuy Lopez ClosedNo ratings yet
- The Joy of Computing Using Python - Unit 2 - Week 0Document3 pagesThe Joy of Computing Using Python - Unit 2 - Week 0Ruy Lopez ClosedNo ratings yet
- Cooperative Cache Management in Mobile Ad Hoc NetworksDocument7 pagesCooperative Cache Management in Mobile Ad Hoc NetworksRuy Lopez ClosedNo ratings yet
- O89n6 T o CDocument303 pagesO89n6 T o CRuy Lopez Closed100% (1)
- 2010 Article 311Document21 pages2010 Article 311Ruy Lopez ClosedNo ratings yet
- Beamer Cheat SheetDocument5 pagesBeamer Cheat SheetRuy Lopez ClosedNo ratings yet
- Gcse Biology Edexcel B1 The Variety of Living Organism Target SheetDocument5 pagesGcse Biology Edexcel B1 The Variety of Living Organism Target SheetSuki ChanNo ratings yet
- Lesson 1Document11 pagesLesson 1remshopNo ratings yet
- Cell AnalogyDocument2 pagesCell AnalogyVea Ann CantollasNo ratings yet
- Long Test g7Document2 pagesLong Test g7Earn cruzNo ratings yet
- Cell Handwritten NotesDocument20 pagesCell Handwritten NotesNavya MehtaNo ratings yet
- Lesson Plan in ScienceDocument5 pagesLesson Plan in ScienceJESPHER GARCIANo ratings yet
- Biology - Exam #2Document14 pagesBiology - Exam #2Juliana RiveraNo ratings yet
- Cell Organelles 2Document10 pagesCell Organelles 2Hardik tyagiNo ratings yet
- Ultra Structure of Plant Cell 1Document18 pagesUltra Structure of Plant Cell 1Kumaran JothiramNo ratings yet
- Workbook Answers PDFDocument36 pagesWorkbook Answers PDFPeriperi PeriperiNo ratings yet
- 8 TH C V Raman English Notes1Document25 pages8 TH C V Raman English Notes1malaisamy43235No ratings yet
- Science: Second Quarter - Module 3B The Plant CellDocument23 pagesScience: Second Quarter - Module 3B The Plant CellRuth LabuacNo ratings yet
- Rajah 1 Di Bawah Menunjukkan Struktur Sejenis Sel.: Bahagian A (20 Markah)Document12 pagesRajah 1 Di Bawah Menunjukkan Struktur Sejenis Sel.: Bahagian A (20 Markah)maysaraNo ratings yet
- Prokaryotic Vs Eukaryotic Cells NotesDocument4 pagesProkaryotic Vs Eukaryotic Cells NotesMischi Jeda ElumbaNo ratings yet
- Biota Quiz Bee Mock Test Part 1Document2 pagesBiota Quiz Bee Mock Test Part 1Ken Juliana Fe IsaacNo ratings yet
- Prokariotik Dan Eukariotik 2Document22 pagesProkariotik Dan Eukariotik 2hanumNo ratings yet
- s.3 Is-Bio Ch.2 WsDocument13 pagess.3 Is-Bio Ch.2 WsChris WongNo ratings yet
- PHA204 Chapter - 1 - 1Document40 pagesPHA204 Chapter - 1 - 1Abdur RaquibNo ratings yet
- Gen Bio 2Document12 pagesGen Bio 2Ma.Dulce ManalastasNo ratings yet
- Click: Cambridge IGCSE Biology Extended LevelDocument20 pagesClick: Cambridge IGCSE Biology Extended Levelkaziba stephenNo ratings yet
- Laboratory Report in Microbiology and ParasitologyDocument3 pagesLaboratory Report in Microbiology and ParasitologyynaNo ratings yet
- Structure and Function of CellsDocument93 pagesStructure and Function of CellsRonald BakangNo ratings yet
- Sankalp Sanjeevni Neet: BiologyDocument13 pagesSankalp Sanjeevni Neet: BiologyKey RavenNo ratings yet
- Parts of A CellDocument16 pagesParts of A CellThomas417No ratings yet
- Selina Solutions For Class 9 Biology Chapter 2 Cell The Unit of LifeDocument15 pagesSelina Solutions For Class 9 Biology Chapter 2 Cell The Unit of LifePrashun PriyadarshiNo ratings yet
- Macmillan Science Level 6 Pupil S Book Unit 1 PP 11 16Document6 pagesMacmillan Science Level 6 Pupil S Book Unit 1 PP 11 16reema2050reemaNo ratings yet