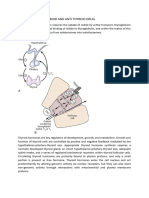
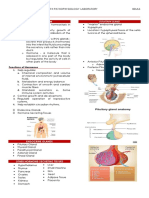
You might also like
- Endocrine Equilibrium: Navigating the Hormonal Seas: Navigating the Whispers of Hormones: A Delicate Dance of BalanceFrom EverandEndocrine Equilibrium: Navigating the Hormonal Seas: Navigating the Whispers of Hormones: A Delicate Dance of BalanceNo ratings yet
- Accorroni Et AlDocument17 pagesAccorroni Et Alp.sepulveda.fiNo ratings yet
- Efectos en El Metabolismo Del Hypo e HipertiroidismoDocument16 pagesEfectos en El Metabolismo Del Hypo e HipertiroidismoLuis Ricardo Hernandez GarciaNo ratings yet
- Analysis of Vitamin C NewDocument59 pagesAnalysis of Vitamin C Newrose mariaNo ratings yet
- The Thyroid and Parathyroid GlandsDocument47 pagesThe Thyroid and Parathyroid GlandsRujha Haniena Ahmad RidzuanNo ratings yet
- Thyroid Gland and Its Rule in Human BodyDocument9 pagesThyroid Gland and Its Rule in Human BodyDrFarah Emad AliNo ratings yet
- Thyroid and Anti Thyroid DrugDocument5 pagesThyroid and Anti Thyroid Drugtamberahul1256No ratings yet
- Tryptophan Metabolism and Gut-Brain HomeostasisDocument23 pagesTryptophan Metabolism and Gut-Brain HomeostasisLuísa OliveiraNo ratings yet
- Genomic and Non-Genomic Mechanisms of Action of Thyroid Hormones and Their Catabolite 3,5-Diiodo-L-Thyronine in MammalsDocument40 pagesGenomic and Non-Genomic Mechanisms of Action of Thyroid Hormones and Their Catabolite 3,5-Diiodo-L-Thyronine in MammalsMarco GiammancoNo ratings yet
- Hormonas TiroideasDocument16 pagesHormonas TiroideasDiva Bruno OviedoNo ratings yet
- Tura KuDocument104 pagesTura KuFitzgerald BillyNo ratings yet
- Overview of Endo Physiology - HandoutDocument19 pagesOverview of Endo Physiology - HandoutVDiesel99No ratings yet
- Thyroid Hormone Transporters Functional and Cinical AplicationsDocument13 pagesThyroid Hormone Transporters Functional and Cinical AplicationsIván LazcanoNo ratings yet
- Artículo de InvestigaciónDocument18 pagesArtículo de InvestigacióngabrielaNo ratings yet
- Fuction Hormons ThyrodsDocument26 pagesFuction Hormons Thyrodsgiselle155204No ratings yet
- Functions of Thyroid Hormones: March 2016Document27 pagesFunctions of Thyroid Hormones: March 2016تقوى اللهNo ratings yet
- Pharmacology of The Endocrine System PDFDocument74 pagesPharmacology of The Endocrine System PDFGlenn Perez71% (7)
- 1 LipidperoxidationDocument8 pages1 LipidperoxidationAgharid Ali HusseinNo ratings yet
- Thyroid Disease in Thyroglobulin ProteinDocument9 pagesThyroid Disease in Thyroglobulin ProteinManolo Hernandez CuellarNo ratings yet
- Physio RespiDocument17 pagesPhysio RespiJenica SorianoNo ratings yet
- VND Ms-Powerpoint&rendition 1Document49 pagesVND Ms-Powerpoint&rendition 1Muhammad SamiNo ratings yet
- Pharmacology of The Endocrine SystemDocument74 pagesPharmacology of The Endocrine SystemarpanabiswassshetyeNo ratings yet
- Laboratory Exercise 13Document18 pagesLaboratory Exercise 13Jessa SuacitoNo ratings yet
- AP-2 Lab Report - Lab 01 The Endrocine SystemDocument15 pagesAP-2 Lab Report - Lab 01 The Endrocine Systemrcdunfee86% (7)
- The Thyroid Gland: Physiology and Pathophysiology: Endocrine Series #6Document16 pagesThe Thyroid Gland: Physiology and Pathophysiology: Endocrine Series #6نذير الشرعبيNo ratings yet
- La Ciencia de Los EsteroidesDocument6 pagesLa Ciencia de Los EsteroidesTATIANA BARRIOS ESCORCIANo ratings yet
- Rhythms in The Endocrine System of Fish: A Review: ArticleDocument34 pagesRhythms in The Endocrine System of Fish: A Review: ArticleBagas Lantip PrakasaNo ratings yet
- 1016MSC T1 2023 Module 5 Tutorial WorksheetsDocument21 pages1016MSC T1 2023 Module 5 Tutorial WorksheetsTina HussainiNo ratings yet
- 337 Pdfsam NMS PhysiologyDocument166 pages337 Pdfsam NMS PhysiologydrpnnreddyNo ratings yet
- The Hypothalamus and Pituitary GlandDocument44 pagesThe Hypothalamus and Pituitary GlandRujha Haniena Ahmad RidzuanNo ratings yet
- Pharmacology: Pharmacology of Endocrine System: Thyroid Hormone& Anti-Thyroid DrugsDocument8 pagesPharmacology: Pharmacology of Endocrine System: Thyroid Hormone& Anti-Thyroid DrugsRaj Nemala Raj NemalaNo ratings yet
- Dawood ThesisDocument50 pagesDawood ThesisDawood khanNo ratings yet
- Endocrine ResonanceDocument51 pagesEndocrine ResonanceEkta ManglaniNo ratings yet
- 10 Diversity of Endocrine System IIDocument23 pages10 Diversity of Endocrine System IIcutethoybaNo ratings yet
- Thyroid GlandDocument30 pagesThyroid GlandahmedkomranNo ratings yet
- Lecture+18 2021+springDocument21 pagesLecture+18 2021+springWael OsmanNo ratings yet
- Endocrine System: University of SurreyDocument13 pagesEndocrine System: University of SurreyqhqhqNo ratings yet
- Thyroid Hormone Synthesis, Secretion, and TransportDocument5 pagesThyroid Hormone Synthesis, Secretion, and TransportAnonymous UTUWFODCEYNo ratings yet
- Metabolic Endocrinology (Week-2)Document8 pagesMetabolic Endocrinology (Week-2)wasimsafdarNo ratings yet
- Endocrinology SupplementDocument85 pagesEndocrinology Supplementfreecouch100% (1)
- Production of Thyroid Hormone and Its ControlDocument2 pagesProduction of Thyroid Hormone and Its ControlDavid YapNo ratings yet
- Drugs Used in Disorders of Endocrine System Ppt. Book (Lectures 1-6)Document467 pagesDrugs Used in Disorders of Endocrine System Ppt. Book (Lectures 1-6)Marc Imhotep Cray, M.D.100% (1)
- Endocrinology Notes: Veterinary PhysiologyDocument16 pagesEndocrinology Notes: Veterinary PhysiologyBrian AllanNo ratings yet
- Thyroid Gland: Ruswana Anwar, Tono DjuwantonoDocument49 pagesThyroid Gland: Ruswana Anwar, Tono Djuwantonoyuyu tuptupNo ratings yet
- Introduction Endocrinal System Physioloogy: Dr. MasikaDocument78 pagesIntroduction Endocrinal System Physioloogy: Dr. MasikaJacob MasikaNo ratings yet
- Chapter 29 - Thyroid Function and Thyroid DrugsDocument21 pagesChapter 29 - Thyroid Function and Thyroid DrugsCapuNo ratings yet
- Endocrine SystemDocument7 pagesEndocrine SystemNicholas FavourNo ratings yet
- Endocrine System NotesDocument6 pagesEndocrine System NotesHannah Grace CorveraNo ratings yet
- Physiology of Thyroid GlandDocument8 pagesPhysiology of Thyroid Glandأياد المشهدانيNo ratings yet
- Antithyroid Drugs by Sejal Khuman Advanced phARMACOLGY 2Document29 pagesAntithyroid Drugs by Sejal Khuman Advanced phARMACOLGY 2Sejal khumanNo ratings yet
- Takenouchi 20151Document13 pagesTakenouchi 20151Angelique R. AltamiranoNo ratings yet
- C C I C 22: Hemical Oordination AND NtegrationDocument13 pagesC C I C 22: Hemical Oordination AND Ntegrationsaihamsitha199_85739No ratings yet
- The Endocrine SystemDocument7 pagesThe Endocrine SystemcystanarisaNo ratings yet
- Reproductive SystemDocument114 pagesReproductive SystembookaccountNo ratings yet
- Hyperthyroidism Cats Part IDocument10 pagesHyperthyroidism Cats Part IFabiola Andrea Abril ParraNo ratings yet
- Thyroid Physiology and Thyroid Function Testing: Angela M. Leung, Gregory A. BrentDocument16 pagesThyroid Physiology and Thyroid Function Testing: Angela M. Leung, Gregory A. BrentAgustinaNo ratings yet
- Thyroid & Parathyroid HormonesDocument53 pagesThyroid & Parathyroid Hormonesrehmanmughal87654No ratings yet
- Ijerph 18 09434Document23 pagesIjerph 18 09434YHOISS SMIHT MUNOZ CERONNo ratings yet
- Cardiovascular and RenalDocument21 pagesCardiovascular and RenalMaria Belen Solis TorresNo ratings yet
- Endocrinology Learning Goals - IPHY 4440Document9 pagesEndocrinology Learning Goals - IPHY 4440Geline Joy D. SamillanoNo ratings yet
- Lipide AnalizeDocument21 pagesLipide AnalizeSandu AlexandraNo ratings yet
- 1 s2.0 S1479666X17300562 MainDocument12 pages1 s2.0 S1479666X17300562 MainSandu AlexandraNo ratings yet
- Joi 150056Document9 pagesJoi 150056Rendy SusantoNo ratings yet
- Poillucci2017 Article LaparoscopicAppendectomyVsAntiDocument10 pagesPoillucci2017 Article LaparoscopicAppendectomyVsAntiSandu AlexandraNo ratings yet
- Allievi2017 Article AcuteAppendicitisStillASurgicaDocument9 pagesAllievi2017 Article AcuteAppendicitisStillASurgicaSandu AlexandraNo ratings yet
- Thyroid Surgery Throughout History: EditorialDocument4 pagesThyroid Surgery Throughout History: EditorialSandu AlexandraNo ratings yet
- 1 s2.0 S1438422106000518 MainDocument5 pages1 s2.0 S1438422106000518 MainSandu AlexandraNo ratings yet
- Butt 2019Document5 pagesButt 2019Sandu AlexandraNo ratings yet
- E00261-16 FullDocument12 pagesE00261-16 FullSandu AlexandraNo ratings yet
- 1 s2.0 S1214021X14600260 MainDocument10 pages1 s2.0 S1214021X14600260 MainSandu AlexandraNo ratings yet
- 1 s2.0 S0196655316001693 MainDocument3 pages1 s2.0 S0196655316001693 MainSandu AlexandraNo ratings yet
- 1 s2.0 S0011502907001034 MainDocument8 pages1 s2.0 S0011502907001034 MainSandu AlexandraNo ratings yet
- 1 s2.0 S1198743X14613203 MainDocument11 pages1 s2.0 S1198743X14613203 MainSandu AlexandraNo ratings yet
- Surgical Techniques For Management Hepatic Hydatid Cysts in Fallujah Teaching Hospital: A Prospective StudyDocument4 pagesSurgical Techniques For Management Hepatic Hydatid Cysts in Fallujah Teaching Hospital: A Prospective StudySandu AlexandraNo ratings yet
- The Current Protocols in The Management of Hepatic Hydatid DiseaseDocument8 pagesThe Current Protocols in The Management of Hepatic Hydatid DiseaseSandu AlexandraNo ratings yet
- Successful Percutaneous Treatment of Extrahepatic Cystic Echinococcosis Through PAIR and Single Puncture Catheter TechniquesDocument7 pagesSuccessful Percutaneous Treatment of Extrahepatic Cystic Echinococcosis Through PAIR and Single Puncture Catheter TechniquesSandu AlexandraNo ratings yet
- Treatment of Intra-Abdominal Hydatid Cyst Using Modified PAIR Technique: Single Center ExperiencesDocument5 pagesTreatment of Intra-Abdominal Hydatid Cyst Using Modified PAIR Technique: Single Center ExperiencesSandu AlexandraNo ratings yet
- Comparison of Long-Term Results of Percutaneous Treatment Techniques For Hepatic Cystic Echinococcosis Types 2 and 3bDocument7 pagesComparison of Long-Term Results of Percutaneous Treatment Techniques For Hepatic Cystic Echinococcosis Types 2 and 3bSandu AlexandraNo ratings yet
- Percutaneous Treatment of Liver Hydatid Cysts in 190 Patients: A Retrospective StudyDocument9 pagesPercutaneous Treatment of Liver Hydatid Cysts in 190 Patients: A Retrospective StudySandu AlexandraNo ratings yet
- Operative Treatment of Hepatic Hydatid Cysts: A Single Center ExperienceDocument6 pagesOperative Treatment of Hepatic Hydatid Cysts: A Single Center ExperienceSandu AlexandraNo ratings yet
- Major Liver Resection For Recurrent Hydatid Cyst of The Liver After Suboptimal TreatmentDocument6 pagesMajor Liver Resection For Recurrent Hydatid Cyst of The Liver After Suboptimal TreatmentSandu AlexandraNo ratings yet
- Arab Journal of GastroenterologyDocument9 pagesArab Journal of GastroenterologySandu AlexandraNo ratings yet
- Hepatic Hydatid Cyst Misdiagnosed As Simple Cyst: A Case ReportDocument6 pagesHepatic Hydatid Cyst Misdiagnosed As Simple Cyst: A Case ReportSandu AlexandraNo ratings yet
- 8Document8 pages8Sandu AlexandraNo ratings yet
- Factors Affecting The Choice of Treatment in Hepatic Hydatid Cyst SurgeryDocument7 pagesFactors Affecting The Choice of Treatment in Hepatic Hydatid Cyst SurgerySandu AlexandraNo ratings yet
- Clinicalinvestigation Non-VascularinterventionsDocument7 pagesClinicalinvestigation Non-VascularinterventionsSandu AlexandraNo ratings yet
- Ovarian Fibromas and Cystadenofibromas: Features of The Fibrous ComponentDocument7 pagesOvarian Fibromas and Cystadenofibromas: Features of The Fibrous ComponentSandu AlexandraNo ratings yet
- Advanced Gynecologic Endoscopy: Edited by Atef DarwishDocument344 pagesAdvanced Gynecologic Endoscopy: Edited by Atef DarwishSandu Alexandra100% (1)
- A Novel Modified PAIR Technique Using A Trocar Catheter For Percutaneous Treatment of Liver Hydatid Cysts: A Six-Year ExperienceDocument5 pagesA Novel Modified PAIR Technique Using A Trocar Catheter For Percutaneous Treatment of Liver Hydatid Cysts: A Six-Year ExperienceSandu AlexandraNo ratings yet
- Case ReportDocument5 pagesCase ReportNurRohmanNo ratings yet
- 2004 - Metabolic and Therapeutic Aspects of Amino Acids in Clinical Nutrition PDFDocument778 pages2004 - Metabolic and Therapeutic Aspects of Amino Acids in Clinical Nutrition PDFlouise kartikaNo ratings yet
- Sodium and PotassiumDocument7 pagesSodium and PotassiumLUALHATI VILLASNo ratings yet
- Ncert - Exemplar - Control and CoordinationDocument18 pagesNcert - Exemplar - Control and CoordinationRagini BaghelNo ratings yet
- ICSE QUESTION PAPER Biology 2010 Class 10 PDFDocument11 pagesICSE QUESTION PAPER Biology 2010 Class 10 PDFLokesh MalikNo ratings yet
- Endocrine System IntroductionDocument5 pagesEndocrine System IntroductionAnnapurna DangetiNo ratings yet
- Unacademy 1Document18 pagesUnacademy 1ANUPAM ANAND KUMAR PANDEY MBA-INo ratings yet
- Christopher D. Moyes, Patricia M. Schulte - Principles of Animal Physiology-Pearson (2016)Document775 pagesChristopher D. Moyes, Patricia M. Schulte - Principles of Animal Physiology-Pearson (2016)salvatore.sara323No ratings yet
- Systems of Intercellaneous Int: 2. The EndocrineDocument62 pagesSystems of Intercellaneous Int: 2. The EndocrineSardar jiNo ratings yet
- Worksheet - I: Biology XDocument5 pagesWorksheet - I: Biology XDharmendra SinghNo ratings yet
- Principle of Animal Physiology (Christopher D. Moyes, Patricia Schulte) Principle ofDocument763 pagesPrinciple of Animal Physiology (Christopher D. Moyes, Patricia Schulte) Principle ofKing Amv88% (8)
- Ana & Physio-NotesDocument82 pagesAna & Physio-NotesAbbie Jane MatigaNo ratings yet
- Approved Curriculam BS-Dental Technology Feb.-2017. 1Document57 pagesApproved Curriculam BS-Dental Technology Feb.-2017. 1Aman WazirNo ratings yet
- CalcitoninDocument2 pagesCalcitoninsandeepab100% (1)
- The Barbell Prescription PDFDocument376 pagesThe Barbell Prescription PDFRINNA100% (2)
- Human Physiology From Cells To Systems Sherwood 9th Edition Test BankDocument15 pagesHuman Physiology From Cells To Systems Sherwood 9th Edition Test Bankandrewmccoymrpazkcnwi100% (48)
- Gen. Pharmacology Tests PDFDocument69 pagesGen. Pharmacology Tests PDFibrahim100% (2)
- Bio f4 Endocrine SystemDocument6 pagesBio f4 Endocrine SystemE-SKUUL ACADEMYNo ratings yet
- Lecture 1, Introduction To EndocrinologyDocument86 pagesLecture 1, Introduction To EndocrinologyAlexandru Ionut Popistaș100% (1)
- fs37 Hormones PDFDocument6 pagesfs37 Hormones PDFadeelyaseen1No ratings yet
- Endocrine Case StudyDocument3 pagesEndocrine Case StudyJenny ViNo ratings yet
- Test Bank Endocrine PDFDocument29 pagesTest Bank Endocrine PDFWenzy Cruz100% (3)
- Questions of PESSI Test For Pharmacists Held On 17 May 2015Document16 pagesQuestions of PESSI Test For Pharmacists Held On 17 May 2015Zirwa AsimNo ratings yet
- Endocrine Exam 1 ReviewDocument11 pagesEndocrine Exam 1 ReviewJohn SmithNo ratings yet
- Assignment Biology Class X Unit - 1Document25 pagesAssignment Biology Class X Unit - 1OjasNo ratings yet
- Hormones and BehaviourDocument16 pagesHormones and BehaviourSushmit KishoreNo ratings yet
- Ix - Nso Set - B Ggn-2Document19 pagesIx - Nso Set - B Ggn-2Priyanka NegiNo ratings yet
- Lesson 4 The Human BrainDocument39 pagesLesson 4 The Human BrainBenNo ratings yet
- Increadible Nervous System PDFDocument48 pagesIncreadible Nervous System PDFmiji_gg100% (1)
- Science f4 Chapter 1 and 2Document4 pagesScience f4 Chapter 1 and 2Halizah RamthanNo ratings yet
- 0610 s18 QP 43Document20 pages0610 s18 QP 43Ray PeramathevanNo ratings yet