You might also like
- Histology EpitheliumDocument18 pagesHistology EpitheliumMarina TeahariNo ratings yet
- Epithelial Tissue PDFDocument20 pagesEpithelial Tissue PDFshafa cahyani100% (1)
- Histology of The Nervous System 2011Document33 pagesHistology of The Nervous System 2011Ng Bing JueNo ratings yet
- Osmosis High-Yield Physiology - Medicalstudyzone - Com (Sürüklenen)Document87 pagesOsmosis High-Yield Physiology - Medicalstudyzone - Com (Sürüklenen)Dilay MercimekNo ratings yet
- Afifi AK, Functional Neuroanatomy. 2005 (Book)Document873 pagesAfifi AK, Functional Neuroanatomy. 2005 (Book)Eguzki Izukaitz NogueraNo ratings yet
- Epithelial TissuesDocument28 pagesEpithelial TissuesHamza SultanNo ratings yet
- Ganong MCQ + Answer PDFDocument72 pagesGanong MCQ + Answer PDFمحمد عبدالله ابراهيم عبدالله81% (16)
- Nervous System Concept MapDocument12 pagesNervous System Concept MapJo BrackenNo ratings yet
- Scientific American Mind, Vol. 14.5 (December 2004)Document101 pagesScientific American Mind, Vol. 14.5 (December 2004)jbmacielNo ratings yet
- Dilek Akakin Epithelial TissueDocument65 pagesDilek Akakin Epithelial TissueErdem AltunNo ratings yet
- Dilek Akakin Epithelial TissueDocument65 pagesDilek Akakin Epithelial TissueErdem AltunNo ratings yet
- HistocologyDocument10 pagesHistocologyKelvin HillsNo ratings yet
- Histology of The Epithelium TissueDocument9 pagesHistology of The Epithelium TissueKelvin HillsNo ratings yet
- Epithelia Tissue: 16 TH January, 2022Document73 pagesEpithelia Tissue: 16 TH January, 2022Anthony KabadiNo ratings yet
- Epithelium: Dr. Kalpajyoti BhattacharjeeDocument125 pagesEpithelium: Dr. Kalpajyoti Bhattacharjeelochi gmNo ratings yet
- Epithelium: Prepared By: Michael John R. Aguilar, RMTDocument124 pagesEpithelium: Prepared By: Michael John R. Aguilar, RMTFrankenstein MelancholyNo ratings yet
- Structural Organisation in AnimalsDocument91 pagesStructural Organisation in AnimalsDivya AgarawalNo ratings yet
- Structural Organisation in AnimalsDocument80 pagesStructural Organisation in AnimalsDivya AgarawalNo ratings yet
- Epithelium (Epithelial Tissue)Document64 pagesEpithelium (Epithelial Tissue)Nihal BilalNo ratings yet
- EPITHELIAL TISSUE (Defn, Classification & Functions - PDFDocument29 pagesEPITHELIAL TISSUE (Defn, Classification & Functions - PDFnikitachaudhary1220No ratings yet
- Epithelial TissueDocument5 pagesEpithelial TissueSofia LiNo ratings yet
- 7.epihelial I MembaranesDocument54 pages7.epihelial I MembaranesRaymond LundaNo ratings yet
- Epithelium: Dr. Bhuvaneswari CDocument23 pagesEpithelium: Dr. Bhuvaneswari CAjinkya AravindNo ratings yet
- محاضره 2 انسجهDocument44 pagesمحاضره 2 انسجهYahya Daham Zafeer SakhrNo ratings yet
- Epithelium: (Edit) Classification (Structural)Document4 pagesEpithelium: (Edit) Classification (Structural)Mirriam Joy CortezNo ratings yet
- L2.1 - Simple Epithelium, 2march 2022Document32 pagesL2.1 - Simple Epithelium, 2march 2022TalarNo ratings yet
- Tob 2.1Document32 pagesTob 2.1LT DRAGONXNo ratings yet
- Epithelial Tissues 1Document87 pagesEpithelial Tissues 1Ayro Business CenterNo ratings yet
- Document 3Document5 pagesDocument 3Chelsea MansuetoNo ratings yet
- Chapter 3 Human Histology - Epithelial TissuesDocument7 pagesChapter 3 Human Histology - Epithelial TissuesDessirie EnriquezNo ratings yet
- Epithelia DR - RaviDocument60 pagesEpithelia DR - Ravi安 娜 胡No ratings yet
- Tissues: Prepared By: Arthur Christian B. Mangio, PTRPDocument80 pagesTissues: Prepared By: Arthur Christian B. Mangio, PTRPjm fabellar100% (1)
- Chapter4 Anaphy+tissuesDocument12 pagesChapter4 Anaphy+tissuesCASSANDRA BARQUILLANo ratings yet
- Ep It HeliumDocument48 pagesEp It HeliumLoisNo ratings yet
- Histology 2018 1Document38 pagesHistology 2018 1Marvin Ndashimye100% (1)
- Epithelial TissueDocument68 pagesEpithelial TissueJASMEEN RAVALNo ratings yet
- Histopathology I: Epithelium Tissue Part IDocument44 pagesHistopathology I: Epithelium Tissue Part ICarinaJongLeeNo ratings yet
- Ep It HeliumDocument75 pagesEp It HeliumSahin mollickNo ratings yet
- TissuesDocument80 pagesTissueskrmgxc8p4fNo ratings yet
- Epithelial TissueDocument68 pagesEpithelial TissueH R A K KulathilakaNo ratings yet
- Elementary TissuesDocument32 pagesElementary TissuesManjunath100% (1)
- Bahasa InggrisDocument5 pagesBahasa Inggrisahmad jamaluddinNo ratings yet
- EpituleinDocument3 pagesEpituleinAlexandra JunchayaNo ratings yet
- Epithelial TissueDocument33 pagesEpithelial TissueStnrhalizaNo ratings yet
- Stratified Epithelium Histology LectureDocument42 pagesStratified Epithelium Histology LectureWajahat FaizNo ratings yet
- Lab SupplementDocument5 pagesLab SupplementcaterappleyeNo ratings yet
- Crissman Epithelial ObjectivesDocument6 pagesCrissman Epithelial Objectiveskkonci01No ratings yet
- محاضره 1 انسجهDocument32 pagesمحاضره 1 انسجهYahya Daham Zafeer SakhrNo ratings yet
- Laboratory ManualDocument39 pagesLaboratory ManualChelsea Chadwick100% (1)
- Histopathology I: Epithelium Tissue Part IIDocument54 pagesHistopathology I: Epithelium Tissue Part IICarinaTingEeNo ratings yet
- MT 100 Lab Notes 1Document11 pagesMT 100 Lab Notes 1Nilo Golosino IIINo ratings yet
- Epithelial TissuesDocument14 pagesEpithelial TissuesJhunelyn100% (2)
- Chapter 3 The EpitheliumDocument3 pagesChapter 3 The EpitheliumGela ReyesNo ratings yet
- Epithelial Tissue Ana205Document30 pagesEpithelial Tissue Ana205Ibrahim BabatundeNo ratings yet
- EpithelialDocument48 pagesEpithelialMbali MazongweNo ratings yet
- Anatomy Lecture NotesDocument94 pagesAnatomy Lecture Notesbiddyusmc100% (2)
- Jaringan EpitelDocument20 pagesJaringan EpitelKitsune TakaNo ratings yet
- Tissue of Body - EpitheliumDocument25 pagesTissue of Body - Epitheliumahmadzaffar8798No ratings yet
- Ciliated Epithelium Is A Category of EpitheliumDocument20 pagesCiliated Epithelium Is A Category of EpitheliumMisnawatiNo ratings yet
- Ana Histology PDFDocument79 pagesAna Histology PDFNagesh NNo ratings yet
- Genbio2 Week 1Document35 pagesGenbio2 Week 1Noel Vincent Agonoy100% (1)
- Stratified Epithelia L 2Document45 pagesStratified Epithelia L 2srzvzvt7rzNo ratings yet
- BIO2OO - Introduction Tissues, Classification of Living Things & Ecology 1.1.0 Animal TissueDocument19 pagesBIO2OO - Introduction Tissues, Classification of Living Things & Ecology 1.1.0 Animal TissueMark SullivanNo ratings yet
- Physiology U-3 Excitable Nervous TissueDocument122 pagesPhysiology U-3 Excitable Nervous Tissuesinte beyuNo ratings yet
- KEY MED 2 Updated 2021 Feb Unit 81Document10 pagesKEY MED 2 Updated 2021 Feb Unit 81huyenhungho5892No ratings yet
- MIDTERM Pilar College Anatomy Physiology Final EditDocument41 pagesMIDTERM Pilar College Anatomy Physiology Final EditYeona BaeNo ratings yet
- LAB EXERCISE4 TISSUES SilveroDocument4 pagesLAB EXERCISE4 TISSUES SilveroGeia Marie SilveroNo ratings yet
- Anatomy and Physiology GMCDocument15 pagesAnatomy and Physiology GMCsamantha tanoNo ratings yet
- Cramer Et Al 2019 Neurologic Medical Device Overview For PathologistsDocument14 pagesCramer Et Al 2019 Neurologic Medical Device Overview For Pathologistsb00403007No ratings yet
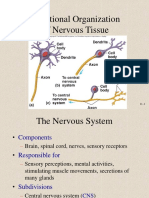
- Functional Organization of Nervous TissueDocument28 pagesFunctional Organization of Nervous TissueIssa JoyahaNo ratings yet
- CH 8 Nervous SystemPractice Test and KeyDocument13 pagesCH 8 Nervous SystemPractice Test and KeyvpcpharmdNo ratings yet
- Ain Aging and Neurodegeneration 1998 EbookDocument267 pagesAin Aging and Neurodegeneration 1998 EbookbebleffNo ratings yet
- Gibco Neurobiology Protocol Handbook Withcover 2021Document136 pagesGibco Neurobiology Protocol Handbook Withcover 2021zikrilchaniagoNo ratings yet
- Terminologia NeuroanatomicaDocument144 pagesTerminologia Neuroanatomicamatteo_vavassoriNo ratings yet
- K8a Anatomi Sistem Saraf PusatDocument87 pagesK8a Anatomi Sistem Saraf Pusathartinissa vaniaNo ratings yet
- Connective TissueDocument202 pagesConnective TissuePeter ThompsonNo ratings yet
- Nervous SystemDocument11 pagesNervous Systemmohd nazmieNo ratings yet
- Lent 2011Document9 pagesLent 2011Marcelo SoaresNo ratings yet
- Introduction To Psychology NotesDocument157 pagesIntroduction To Psychology NotesIt's sean bwoy 7No ratings yet
- UNIT 2 Lesson 2.2 Structures of Animal CellsDocument24 pagesUNIT 2 Lesson 2.2 Structures of Animal CellsKatznjammr100% (1)
- Multiple Choice QuizDocument7 pagesMultiple Choice QuizNargess OsmanNo ratings yet
- BIO 35 Chapter 2 - Nerve PhysiologyDocument14 pagesBIO 35 Chapter 2 - Nerve PhysiologyJake EverettNo ratings yet
- Chapter 20 - MolluscaGastropoda CompressedDocument27 pagesChapter 20 - MolluscaGastropoda Compressedaulia rahmahNo ratings yet
- Termin SarafDocument26 pagesTermin SarafSyifa KaniaNo ratings yet
- Neurogenesis and Maturation I Brain InjuryDocument11 pagesNeurogenesis and Maturation I Brain Injuryseemee23820No ratings yet
- A&P Chapter 8 Nervous SystemDocument32 pagesA&P Chapter 8 Nervous SystemJeong song park100% (1)
- A Glossary: of Key Brain Science TermsDocument24 pagesA Glossary: of Key Brain Science TermscadulemosNo ratings yet