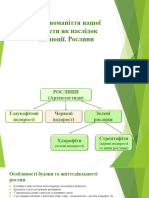
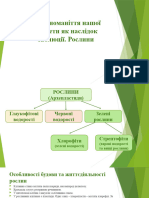
You might also like
- Залежність ростових процесів від різних факторівDocument16 pagesЗалежність ростових процесів від різних факторівАнастасія МикитюкNo ratings yet
- L 19-2Document18 pagesL 19-2viviNo ratings yet
- Залежність ростових процесів від різних факторівDocument16 pagesЗалежність ростових процесів від різних факторівАнастасія МикитюкNo ratings yet
- Вплив Строків Сівби Різних Сортів Озимого Ячменю ТаDocument10 pagesВплив Строків Сівби Різних Сортів Озимого Ячменю ТаИринаNo ratings yet
- Реферат біотехнологіїDocument12 pagesРеферат біотехнологіїВікторія ПроскураNo ratings yet
- Алелопатія лекціяDocument16 pagesАлелопатія лекціяЛюбов100% (1)
- 14779-Article Text-29217-1-10-20191207Document12 pages14779-Article Text-29217-1-10-20191207julia chuharevaNo ratings yet
- ФітогормониDocument9 pagesФітогормониgamemax2007lixNo ratings yet
- Урок 21 Автотрофні та гетеротрофні організмиDocument11 pagesУрок 21 Автотрофні та гетеротрофні організмиinaNo ratings yet
- Лекція Розвиток рослинDocument6 pagesЛекція Розвиток рослинoeugenneNo ratings yet
- реферат зкксDocument6 pagesреферат зкксSound MeNo ratings yet
- 11 Основні с-ща існуванняDocument18 pages11 Основні с-ща існуванняДаша ЗадорожнаNo ratings yet
- Attach 15281378878200Document32 pagesAttach 15281378878200Galya StarchenkoNo ratings yet
- Attach 15281378878200Document32 pagesAttach 15281378878200Galya StarchenkoNo ratings yet
- Лекція 3Document24 pagesЛекція 3nperederiyNo ratings yet
- фотосинтезDocument25 pagesфотосинтезVadimNo ratings yet
- ліс - легені нашої планетиDocument21 pagesліс - легені нашої планетиРуслан СавчукNo ratings yet
- Upload 1370573 1711961087833673504Document3 pagesUpload 1370573 1711961087833673504ingaodessa100% (1)
- Ркð Ð Ð Ð Ñ Ñ Ð Ñ Ñ Ð°ðºñ Ð Ñ Ð 1Document34 pagesРкð Ð Ð Ð Ñ Ñ Ð Ñ Ñ Ð°ðºñ Ð Ñ Ð 1appolinariya767No ratings yet
- Вирощування Органiчноi Продукцii з Застосуванням Органiчних ДобривDocument46 pagesВирощування Органiчноi Продукцii з Застосуванням Органiчних ДобривtanyaNo ratings yet
- Адаптація Рослин До Високих ТемпературDocument5 pagesАдаптація Рослин До Високих ТемпературLoove Sleep67% (3)
- практична робота №2Document4 pagesпрактична робота №2Ирина ВолокитинаNo ratings yet
- Лекція 22 ФітогормониDocument33 pagesЛекція 22 ФітогормониАнастасія МикитюкNo ratings yet
- Адаптивні біологічні ритмиDocument5 pagesАдаптивні біологічні ритмиЕгор ОняновNo ratings yet
- Будова та форми клітин справжніх бактерійDocument4 pagesБудова та форми клітин справжніх бактерійАльбіна БасNo ratings yet
- Диво фотосинтезу»Document8 pagesДиво фотосинтезу»Фазекаш ДмитроNo ratings yet
- Диво фотосинтезу»Document8 pagesДиво фотосинтезу»Фазекаш ДмитроNo ratings yet
- Презентація 8Document7 pagesПрезентація 8сайбірдодя77No ratings yet
- Б-ОТЕХНОЛОГ-Я В¦там¦ни як складов¦ компоненти поживних середовищDocument12 pagesБ-ОТЕХНОЛОГ-Я В¦там¦ни як складов¦ компоненти поживних середовищЛюбовNo ratings yet
- Урок 52 Екосистеми. Ланцюги живлення.Document7 pagesУрок 52 Екосистеми. Ланцюги живлення.FleazzyNo ratings yet
- 207432Document35 pages207432Дарья ЛуценкоNo ratings yet
- Лекція1. ВступDocument7 pagesЛекція1. Вступviktoriia.dzhus.21No ratings yet
- UntitledDocument21 pagesUntitledСтанислав КоноваленкоNo ratings yet
- Як розвиваються рослиниDocument2 pagesЯк розвиваються рослиниkotyalipsikNo ratings yet
- Nanotehnologiyi V Suchasnomu Silskomu GospodarstviDocument4 pagesNanotehnologiyi V Suchasnomu Silskomu GospodarstviУляна ТютькоNo ratings yet
- біогазDocument14 pagesбіогазОксана ГудзьNo ratings yet
- 20090Document9 pages20090Діана ЮручукNo ratings yet
- 10 Клас Урок 14 Водорості.Document11 pages10 Клас Урок 14 Водорості.asalexanderbalabasssNo ratings yet
- ЦитокініниDocument24 pagesЦитокініниАнастасія МикитюкNo ratings yet
- Екологія Псамофітів і ГелофітівDocument15 pagesЕкологія Псамофітів і ГелофітівОлег РосущанNo ratings yet
- ЕкосистемаDocument4 pagesЕкосистемаСоломія МілянNo ratings yet
- презентація 13Document19 pagesпрезентація 13АняNo ratings yet
- Вступ до фаху 6 рDocument7 pagesВступ до фаху 6 рsergey24 t.No ratings yet
- обмін речовинDocument19 pagesобмін речовинНіна ДабіжаNo ratings yet
- 10 Клас Урок 14 Водорості.Document11 pages10 Клас Урок 14 Водорості.asalexanderbalabasssNo ratings yet
- Бешлей 2011 ФітотестуванняDocument3 pagesБешлей 2011 ФітотестуванняYaroslavNo ratings yet
- лекція 2 - частина 1Document16 pagesлекція 2 - частина 1Світлана ГуменнаNo ratings yet
- UntitledDocument21 pagesUntitledСтанислав КоноваленкоNo ratings yet
- 1Document4 pages1Лиза КовалюкNo ratings yet
- біологіяDocument2 pagesбіологіяІлля ФещукNo ratings yet
- ZitologiaDocument31 pagesZitologiaKirill PashkevichNo ratings yet
- Уроки 3-4 11 класDocument8 pagesУроки 3-4 11 класLerusik DuNo ratings yet
- 327834Document11 pages327834lidllozNo ratings yet
- Біологія -С-21 -25 03Document15 pagesБіологія -С-21 -25 03Margarita DzhamNo ratings yet
- 327834Document10 pages327834dmutro0211nakNo ratings yet
- UntitledDocument5 pagesUntitledАнастасия КосюгNo ratings yet
- рефератDocument16 pagesрефератОксана СловачевськаNo ratings yet
- 1. БессоноваDocument5 pages1. Бессоноваcrazyproff1911No ratings yet
- ЫЫЫЫыDocument10 pagesЫЫЫЫыsetroninNo ratings yet
- UntitledDocument536 pagesUntitledАндрій ФедоровNo ratings yet
- Kovalenko O.RZ Roslinnitstvo Lab 2014Document140 pagesKovalenko O.RZ Roslinnitstvo Lab 2014Андрій ФедоровNo ratings yet
- 21-22 MergedDocument41 pages21-22 MergedАндрій ФедоровNo ratings yet
- Селекція сг культур - ЛЗ - 12Document2 pagesСелекція сг культур - ЛЗ - 12Андрій ФедоровNo ratings yet