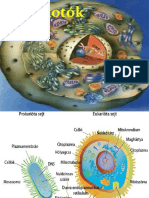
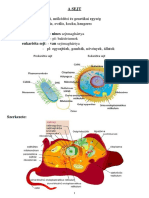
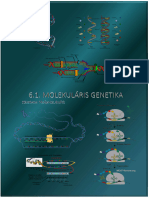
You might also like
- Biológia IIIDocument140 pagesBiológia IIIMazel TovNo ratings yet
- 6.-Kurzus EmbriológiaDocument32 pages6.-Kurzus EmbriológiaNagy MárkNo ratings yet
- Sejtalkotok JDocument50 pagesSejtalkotok Jangelika6ill6s100% (1)
- Daganatképződés. FejezetDocument7 pagesDaganatképződés. FejezetBlazsNo ratings yet
- Daganatképződés. FejezetDocument7 pagesDaganatképződés. FejezetBlazsNo ratings yet
- DR Papp Csaba Fogamzas Es Elettani TerhessegDocument90 pagesDR Papp Csaba Fogamzas Es Elettani Terhesseghppp555No ratings yet
- Fejlődéstan Mi Tételsorunk KidolgozvaDocument98 pagesFejlődéstan Mi Tételsorunk KidolgozvaZsofiaNo ratings yet
- 25.az Egyedfejlődés SzabályozásaDocument10 pages25.az Egyedfejlődés SzabályozásaBlazsNo ratings yet
- Endomembranok BioszferaDocument6 pagesEndomembranok BioszferaBéla KamuNo ratings yet
- Családfa GenetikaDocument8 pagesCsaládfa GenetikaAdrianFenyvesyNo ratings yet
- Egyedfejlodes FaktDocument30 pagesEgyedfejlodes FaktdoroginédNo ratings yet
- Gollesz Viktor FejldstanDocument35 pagesGollesz Viktor FejldstanRebekaNo ratings yet
- TételDocument5 pagesTételViki100% (1)
- TelegóniaDocument1 pageTelegóniaultraboxNo ratings yet
- 09.01.A SejtDocument8 pages09.01.A SejtMolnár GyulaNo ratings yet
- Genetika1 FaktDocument38 pagesGenetika1 Faktanita_nemesNo ratings yet
- Biológia Távoktatási Segédanyag 11evf TeljesDocument11 pagesBiológia Távoktatási Segédanyag 11evf TeljesRebeka MolnárNo ratings yet
- A DNS Örökítő SzerepeDocument3 pagesA DNS Örökítő SzerepebcsmNo ratings yet
- Klónozás (Zsuzsival Közös Munka)Document8 pagesKlónozás (Zsuzsival Közös Munka)peter_soós100% (1)
- Genomic Imprinting - Metiláció, Heterokromatin Gátlás És A GénkonverzióDocument14 pagesGenomic Imprinting - Metiláció, Heterokromatin Gátlás És A GénkonverzióBárány JózsefNo ratings yet
- BIOLÓGIA SEJT SZAPORODÁS Kép NélkülDocument3 pagesBIOLÓGIA SEJT SZAPORODÁS Kép NélkülLilla ForróNo ratings yet
- Altalanos Fejlodestan 2020Document49 pagesAltalanos Fejlodestan 2020Levente SchmidtNo ratings yet
- Sass Miklós: Összehasonlító EmbriológiaDocument7 pagesSass Miklós: Összehasonlító Embriológiagnadori100% (1)
- SzabadJanos A MuslicaDocument16 pagesSzabadJanos A Muslicahelgaa.balogNo ratings yet
- FejlődéstanDocument58 pagesFejlődéstanghjuditNo ratings yet
- A Sejt Genetikája ÚJ Hi-S MásolataDocument5 pagesA Sejt Genetikája ÚJ Hi-S Másolataybd6fc86xcNo ratings yet
- SejtosztodasDocument36 pagesSejtosztodasangelika6ill6sNo ratings yet
- Falus EpigenetikaDocument12 pagesFalus EpigenetikaGéza NagyNo ratings yet
- 2.az Emberi ÖrökségDocument3 pages2.az Emberi ÖrökségSuha ViktóriaNo ratings yet
- Genetika2 FaktDocument24 pagesGenetika2 FaktÁdám PallosNo ratings yet
- Eukarióta És Prokarióta Sejt ÖsszehasonlításaDocument2 pagesEukarióta És Prokarióta Sejt ÖsszehasonlításaFanni KissNo ratings yet
- 11 Előadas-26052017Document70 pages11 Előadas-26052017kereklovagNo ratings yet
- Göllesz Viktor FejlődéstanDocument6 pagesGöllesz Viktor FejlődéstanHajdu-Mustos BernadettNo ratings yet
- Genetika-T Telkidolgoz SDocument25 pagesGenetika-T Telkidolgoz SFulop NoemiNo ratings yet
- Sejtbiológia Feladatlap Márc. 1.Document2 pagesSejtbiológia Feladatlap Márc. 1.Perneki DoraNo ratings yet
- Humángenetika HoffmannDocument30 pagesHumángenetika HoffmannBalogh VivienNo ratings yet
- 6.1 SejtmagDocument62 pages6.1 Sejtmagvargareka47No ratings yet
- 02 PlasztiszokDocument8 pages02 PlasztiszoknemfogomNo ratings yet
- Sejt - És SzövettenyészetekDocument13 pagesSejt - És SzövettenyészetekAlexandra SzőcsNo ratings yet
- Fejlodestan Fogalmak ABCDocument16 pagesFejlodestan Fogalmak ABCdoroginédNo ratings yet
- KlonozasDocument1 pageKlonozasDániel NagyNo ratings yet
- Molekularis GenetikaDocument22 pagesMolekularis GenetikaAnanda VédaNo ratings yet
- Az Eukarióta Sejt - 2Document20 pagesAz Eukarióta Sejt - 22025d Varró AnnaNo ratings yet
- A SejtDocument6 pagesA SejtGyűjtő SzidiNo ratings yet
- Genetika Legujabb 2017Document7 pagesGenetika Legujabb 2017Nagy AnnaNo ratings yet
- CsumiDocument4 pagesCsumimakkai.oliverNo ratings yet
- ProkariótákDocument9 pagesProkariótákdNo ratings yet
- Az Állatok SzaporodásbiológiájaDocument44 pagesAz Állatok SzaporodásbiológiájaManhertz PéterNo ratings yet
- Genetika1 FaktDocument41 pagesGenetika1 FaktSimon ViktóriaNo ratings yet
- Virág 1Document25 pagesVirág 1Dóri PappNo ratings yet
- 7.1. Golgi-Keszulek LizoszomaDocument17 pages7.1. Golgi-Keszulek Lizoszomavargareka47No ratings yet
- Embrionalis FejlodesDocument36 pagesEmbrionalis FejlodesErika EszterNo ratings yet
- Mutációk PrezentációDocument12 pagesMutációk PrezentációKartal DienesNo ratings yet
- GéntechnológiaDocument2 pagesGéntechnológiaPuRe Sp3ctreNo ratings yet
- Sejtbiológia Tételek!Document2 pagesSejtbiológia Tételek!Alexandra FarkasNo ratings yet
- Prion Oláh MDocument11 pagesPrion Oláh MMózesNo ratings yet
- Megtermékenyà - Tã©s és beágyazà Dã¡sDocument16 pagesMegtermékenyà - Tã©s és beágyazà Dã¡sSzekeres-Kovács PetraNo ratings yet
- Anatómia Ea6Document43 pagesAnatómia Ea6Óváry- Papp RékaNo ratings yet
- Subiecte Olimpiada Națională de Biologie Proba Teoretică Clasa A IX-aDocument18 pagesSubiecte Olimpiada Națională de Biologie Proba Teoretică Clasa A IX-aRoxanaGaraimanNo ratings yet
- Előadás 13Document5 pagesElőadás 13BlazsNo ratings yet
- SejtbiohazikDocument33 pagesSejtbiohazikBlazsNo ratings yet
- Komplex Biologia I Gyakorlati Jegyzet 2012Document27 pagesKomplex Biologia I Gyakorlati Jegyzet 2012BlazsNo ratings yet
- Előadás 1Document5 pagesElőadás 1BlazsNo ratings yet
- Adbáz 2Document71 pagesAdbáz 2BlazsNo ratings yet
- Adbáz 1Document27 pagesAdbáz 1BlazsNo ratings yet
- Sejtciklus. FejezetDocument9 pagesSejtciklus. FejezetBlazsNo ratings yet
- Sejtciklus. FejezetDocument9 pagesSejtciklus. FejezetBlazsNo ratings yet
- JelátvitelDocument8 pagesJelátvitelBlazsNo ratings yet
- 14 Génexpresszió ProkariótákbanDocument6 pages14 Génexpresszió ProkariótákbanBlazsNo ratings yet
- Replikáció És Sejtosztódás. FejezetDocument8 pagesReplikáció És Sejtosztódás. FejezetBlazsNo ratings yet