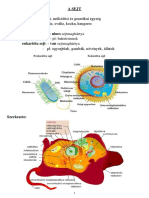
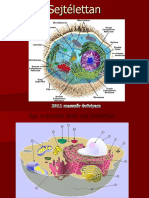
You might also like
- Az Eukarióta Sejt - 2Document20 pagesAz Eukarióta Sejt - 22025d Varró AnnaNo ratings yet
- Altalanos Fejlodestan 2020Document49 pagesAltalanos Fejlodestan 2020Levente SchmidtNo ratings yet
- BIOLÓGIA SEJT SZAPORODÁS Kép NélkülDocument3 pagesBIOLÓGIA SEJT SZAPORODÁS Kép NélkülLilla ForróNo ratings yet
- A Sejt Genetikája ÚJ Hi-S MásolataDocument5 pagesA Sejt Genetikája ÚJ Hi-S Másolataybd6fc86xcNo ratings yet
- About The CellDocument108 pagesAbout The CellDomokos ErzsebetNo ratings yet
- A SejtosztódásDocument22 pagesA SejtosztódásCsenge VajdaNo ratings yet
- Genetika1 FaktDocument38 pagesGenetika1 Faktanita_nemesNo ratings yet
- PDF DokumentumDocument36 pagesPDF DokumentumAdri GollNo ratings yet
- A SejtciklusDocument2 pagesA SejtciklusklaudiaNo ratings yet
- Biológia A Kromoszómák Vizsgálata, Kromoszomális RendellenességekDocument39 pagesBiológia A Kromoszómák Vizsgálata, Kromoszomális RendellenességekKósa AndreaNo ratings yet
- 7 Előadás-28042017cDocument48 pages7 Előadás-28042017ckereklovagNo ratings yet
- 2.A sejt membránnal nem rendelkezőDocument5 pages2.A sejt membránnal nem rendelkezőHorváth AdriennNo ratings yet
- SejtosztodasDocument36 pagesSejtosztodasangelika6ill6sNo ratings yet
- 14. Alapfogalmak_Az öröklődő információ megjelenésének kémiai alapjaiDocument22 pages14. Alapfogalmak_Az öröklődő információ megjelenésének kémiai alapjaiweoresbiologiaNo ratings yet
- Mitózis Meiozis TZsDocument28 pagesMitózis Meiozis TZsPanniNo ratings yet
- Sejt GenetikájaDocument2 pagesSejt Genetikájaanettka19985No ratings yet
- Génexpresszió És Az Azzal Kapcsolatos SejtorganellumokDocument55 pagesGénexpresszió És Az Azzal Kapcsolatos SejtorganellumokNoémi TamaskaNo ratings yet
- Biol 7 10.klik036622001#283937#Tanulo#161063#CsatolmanyDocument1 pageBiol 7 10.klik036622001#283937#Tanulo#161063#CsatolmanyEmma TörökNo ratings yet
- ProkariótákDocument9 pagesProkariótákdNo ratings yet
- 11 - 1 - Bevezetés A Sejtbiológia És A Genetika Világába - 19 - 20 - 2Document35 pages11 - 1 - Bevezetés A Sejtbiológia És A Genetika Világába - 19 - 20 - 2Péter OlláriNo ratings yet
- SpermiumképzésDocument3 pagesSpermiumképzésbe koNo ratings yet
- Férfi Nemi SzervekDocument59 pagesFérfi Nemi SzervekBrigitta MózesNo ratings yet
- 09.01.A SejtDocument8 pages09.01.A SejtMolnár GyulaNo ratings yet
- Genetika1 FaktDocument41 pagesGenetika1 FaktSimon ViktóriaNo ratings yet
- DR Toth Zsuzsanna Mitozis MeiozisDocument28 pagesDR Toth Zsuzsanna Mitozis MeiozismagdiNo ratings yet
- Genetika-T Telkidolgoz SDocument25 pagesGenetika-T Telkidolgoz SFulop NoemiNo ratings yet
- 6.1 SejtmagDocument62 pages6.1 Sejtmagvargareka47No ratings yet
- Meiózis Profázis I Szakaszai MagyarulDocument3 pagesMeiózis Profázis I Szakaszai Magyarulgretus9415No ratings yet
- Genetikafogalmak BiológiaDocument4 pagesGenetikafogalmak BiológiaZak DannyNo ratings yet
- Sejttan 2011Document55 pagesSejttan 2011Brigitta KondrátNo ratings yet
- Humángenetika HoffmannDocument30 pagesHumángenetika HoffmannBalogh VivienNo ratings yet
- Fejlodestan Fogalmak ABCDocument16 pagesFejlodestan Fogalmak ABCdoroginédNo ratings yet
- Göllesz Viktor FejlődéstanDocument6 pagesGöllesz Viktor FejlődéstanHajdu-Mustos BernadettNo ratings yet
- TételDocument5 pagesTételViki100% (1)
- örökítőanyag.fogalmakDocument2 pagesörökítőanyag.fogalmakgtknjshh7hNo ratings yet
- SzabadJanos_A_muslica (1)Document16 pagesSzabadJanos_A_muslica (1)helgaa.balogNo ratings yet
- A Novenyi SejtDocument83 pagesA Novenyi SejtKitti Sztyehlik-OcztosNo ratings yet
- Bevbiosz 2Document7 pagesBevbiosz 2algamark777No ratings yet
- Biológia Távoktatási Segédanyag 11evf TeljesDocument11 pagesBiológia Távoktatási Segédanyag 11evf TeljesRebeka MolnárNo ratings yet
- 1 SejtbioszDocument39 pages1 SejtbioszbolcsbekaNo ratings yet
- Családfa GenetikaDocument8 pagesCsaládfa GenetikaAdrianFenyvesyNo ratings yet
- Eukarióta Egysejtűek - TáblázatDocument2 pagesEukarióta Egysejtűek - TáblázatszabolcsNo ratings yet
- Az Antibiotikum Termeés Genetikai Alapjai-Zsigmond OrsolyaDocument16 pagesAz Antibiotikum Termeés Genetikai Alapjai-Zsigmond Orsolyazsigmondorsolya1No ratings yet
- Nem-Mendeli Genetika. FejezetDocument6 pagesNem-Mendeli Genetika. FejezetBlazsNo ratings yet
- Az Élőlények Szerveződési SzintjeiDocument4 pagesAz Élőlények Szerveződési SzintjeiDani KukudaNo ratings yet
- SodaPDF Merged Merging ResultDocument364 pagesSodaPDF Merged Merging ResultintifintyNo ratings yet
- Genetika Alapjai (2007, 99 Oldal)Document99 pagesGenetika Alapjai (2007, 99 Oldal)Attila TóthNo ratings yet
- Állatgenetika 2024.docxDocument19 pagesÁllatgenetika 2024.docxEllieNo ratings yet
- Eloadas Botanika (Kert. - T.Ep.) 2020-2021Document121 pagesEloadas Botanika (Kert. - T.Ep.) 2020-2021Domokos ErzsebetNo ratings yet
- Tétel SejtbioszDocument23 pagesTétel SejtbioszZsóciNo ratings yet
- Sejbiologia3 5 2009Document44 pagesSejbiologia3 5 2009szecska5No ratings yet
- Sass Miklós: Összehasonlító EmbriológiaDocument7 pagesSass Miklós: Összehasonlító Embriológiagnadori100% (1)
- Genetika2 FaktDocument24 pagesGenetika2 FaktÁdám PallosNo ratings yet
- Az Ember Szaporodása És EgyedfejlődéseDocument11 pagesAz Ember Szaporodása És EgyedfejlődéseLibi TibarovskyNo ratings yet
- JegyzetDocument9 pagesJegyzetintifintyNo ratings yet
- Sejttan JegyzetDocument2 pagesSejttan JegyzetGéza KissNo ratings yet
- 13Document56 pages13Ági HorváthNo ratings yet
- 10 Előadás-19052017aDocument80 pages10 Előadás-19052017akereklovagNo ratings yet
- Biologia Felveteli Tetelek IV 1999Document11 pagesBiologia Felveteli Tetelek IV 1999Károly PappNo ratings yet
- Urologia TortenelemDocument28 pagesUrologia TortenelemkereklovagNo ratings yet
- 3.a.hugyuti InfekciokDocument19 pages3.a.hugyuti InfekciokkereklovagNo ratings yet
- UrográfiaDocument2 pagesUrográfiakereklovagNo ratings yet
- P5 Vese BetegségeiDocument87 pagesP5 Vese BetegségeikereklovagNo ratings yet
- Ossz PPT MagyarDocument593 pagesOssz PPT MagyarkereklovagNo ratings yet
- Eloadas 9 MajDocument44 pagesEloadas 9 MajkereklovagNo ratings yet
- Konyv PDFDocument122 pagesKonyv PDFkereklovagNo ratings yet
- 1.fejlodesi Rendellenessegek PDFDocument50 pages1.fejlodesi Rendellenessegek PDFkereklovagNo ratings yet
- Szuleszeti Es Nogyogyaszati PatologiaDocument91 pagesSzuleszeti Es Nogyogyaszati PatologiakereklovagNo ratings yet
- Ultrahang - I. ElőadásDocument14 pagesUltrahang - I. ElőadáskereklovagNo ratings yet
- Eloadas 13 Ovarium PDFDocument15 pagesEloadas 13 Ovarium PDFkereklovagNo ratings yet
- 4 Előadás-CDocument63 pages4 Előadás-CkereklovagNo ratings yet
- Eloadas 12 Noi Nemi SzervekDocument28 pagesEloadas 12 Noi Nemi SzervekkereklovagNo ratings yet
- Eloadas 8 Gyomor-BelekDocument145 pagesEloadas 8 Gyomor-BelekkereklovagNo ratings yet
- Eloadas 7 LegutakDocument106 pagesEloadas 7 LegutakkereklovagNo ratings yet
- Vizkelety o PDFDocument264 pagesVizkelety o PDFkereklovag67% (3)
- MorfopatologiaDocument32 pagesMorfopatologiakereklovagNo ratings yet
- 7 Előadás-28042017cDocument48 pages7 Előadás-28042017ckereklovagNo ratings yet
- 10 Előadás-19052017aDocument80 pages10 Előadás-19052017akereklovagNo ratings yet
- 7 Előadás-28042017cDocument48 pages7 Előadás-28042017ckereklovagNo ratings yet
- ZsiranyagcsereDocument67 pagesZsiranyagcserekereklovagNo ratings yet
- Bara Tivadar Fizikai Vizsgalat Es Sebeszeti Alapfogalmak 1 PDFDocument171 pagesBara Tivadar Fizikai Vizsgalat Es Sebeszeti Alapfogalmak 1 PDFkereklovagNo ratings yet
- 8 Előadás-05052017bDocument53 pages8 Előadás-05052017bkereklovagNo ratings yet
- SejtbioDocument76 pagesSejtbiokereklovagNo ratings yet
- Urologia PDFDocument268 pagesUrologia PDFAnonymous p28Tktgl5No ratings yet
- Sejtbiológia 1Document57 pagesSejtbiológia 1kereklovagNo ratings yet
- AminoglokozidokDocument51 pagesAminoglokozidokkereklovagNo ratings yet
- SedatohipnotikumokDocument24 pagesSedatohipnotikumokkereklovagNo ratings yet
- 1 Eloadas Kardiovaszkularis Anmanezis, TunetekDocument133 pages1 Eloadas Kardiovaszkularis Anmanezis, TunetekkereklovagNo ratings yet