You might also like
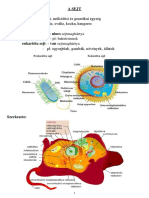
- 09.01.A SejtDocument8 pages09.01.A SejtMolnár GyulaNo ratings yet
- Alapszakdolgozatok, Mesterszakdolgozatok, Diplomamunkák, Szakdolgozatok, Tudományos Megírása Könnyedén (Tudományos Munka)From EverandAlapszakdolgozatok, Mesterszakdolgozatok, Diplomamunkák, Szakdolgozatok, Tudományos Megírása Könnyedén (Tudományos Munka)No ratings yet
- Vér Kórélettana PDFDocument38 pagesVér Kórélettana PDFNikolett NagyNo ratings yet
- Az IdegrendszerDocument57 pagesAz IdegrendszerZsuzsanna Franciska100% (2)
- Korneth Aniko Klinikai Ismeretek A Belgyogyaszat Koreből Apolonők Reszere - 13Document5 pagesKorneth Aniko Klinikai Ismeretek A Belgyogyaszat Koreből Apolonők Reszere - 13ama001No ratings yet
- Reszl KortanDocument55 pagesReszl KortanZoli KünsztlerNo ratings yet
- éLETTAN Nyirok Immunolã Gia - 10Document50 pageséLETTAN Nyirok Immunolã Gia - 10KataNo ratings yet
- A Vér ÉlettanaDocument94 pagesA Vér Élettanakitti2001No ratings yet
- Sav - Bazis Egyensuly - Gazometria - 20190920Document11 pagesSav - Bazis Egyensuly - Gazometria - 20190920Viktória Németh100% (1)
- Reprostanol JóDocument2 pagesReprostanol JósefdeniNo ratings yet
- Az ImmunrendszerDocument66 pagesAz ImmunrendszerRenáta FerencziNo ratings yet
- Korneth Aniko Klinikai Ismeretek A Belgyogyaszat Koreből Apolonők Reszere - 15Document5 pagesKorneth Aniko Klinikai Ismeretek A Belgyogyaszat Koreből Apolonők Reszere - 15ama001No ratings yet
- 05 Neurologia ArcfajdalmakDocument26 pages05 Neurologia ArcfajdalmakHalász MátéNo ratings yet
- Biofizika 2Document12 pagesBiofizika 2Fanni SzabóNo ratings yet
- Rextra 2014 OnlineDocument148 pagesRextra 2014 OnlineTamásKureczNo ratings yet
- XXX. Évfolyam 2022. NyárDocument100 pagesXXX. Évfolyam 2022. NyárVersmondó100% (1)
- Komsa Fruzsina Tóth Miklós - Szervetlen Kémia. Összefoglalás Orvostanhallgatók Számára WWW - Semmelweiskiado.hu (Trid:44031)Document28 pagesKomsa Fruzsina Tóth Miklós - Szervetlen Kémia. Összefoglalás Orvostanhallgatók Számára WWW - Semmelweiskiado.hu (Trid:44031)Fruzsina Eniko KomsaNo ratings yet
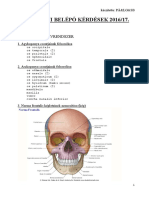
- Szigorlati Belépő Kérdések 2016 1.PDF MásolataDocument55 pagesSzigorlati Belépő Kérdések 2016 1.PDF MásolataNoémi CsehNo ratings yet
- Ápolástani Alapismeretekrev2Document207 pagesÁpolástani Alapismeretekrev2barbi.flora2003No ratings yet
- Korneth Aniko Klinikai Ismeretek A Belgyogyaszat Koreből Apolonők Reszere - 20Document5 pagesKorneth Aniko Klinikai Ismeretek A Belgyogyaszat Koreből Apolonők Reszere - 20ama001No ratings yet
- ChameleonDocument181 pagesChameleonRekaNo ratings yet
- Anatómia Tételek VizsgáraDocument31 pagesAnatómia Tételek VizsgáraokanyirekaNo ratings yet
- Élettanalap JegyzetDocument345 pagesÉlettanalap JegyzetdugoNo ratings yet
- 7 Előadás-28042017cDocument48 pages7 Előadás-28042017ckereklovagNo ratings yet
- Élettan Kórélettan PF12ÉK312Document2 pagesÉlettan Kórélettan PF12ÉK312Hajnalka SzolykaNo ratings yet
- Ipari Mikrobiologia II IIIDocument232 pagesIpari Mikrobiologia II IIISzabó TamásNo ratings yet
- II 02 Nyirokrendszer 02 2014Document12 pagesII 02 Nyirokrendszer 02 2014dorsolya92No ratings yet
- Testfolyadekok, Ver, Szovetnedv, NyirokDocument33 pagesTestfolyadekok, Ver, Szovetnedv, NyirokKbsv1389No ratings yet
- Akut Belgyógyászati EsetekDocument6 pagesAkut Belgyógyászati EsetekBertamonika76No ratings yet
- 4 Előadás-CDocument63 pages4 Előadás-CkereklovagNo ratings yet
- 1 Tétel IdegszövetDocument3 pages1 Tétel IdegszövetAnonymous ZjQ5fUcPNo ratings yet
- Keresztrejtveny Orvosi Latin 28 oDocument2 pagesKeresztrejtveny Orvosi Latin 28 oPetra Andrea GyarmatiNo ratings yet
- Betegségek Klinikuma2019Document12 pagesBetegségek Klinikuma2019ÉnCsakNo ratings yet
- Bevezetes A Makroszkopos AnatomiábaDocument66 pagesBevezetes A Makroszkopos AnatomiábaMolnár EdinaNo ratings yet
- 001 Praktikum 1 2019 20Document191 pages001 Praktikum 1 2019 20tamas.emeseNo ratings yet
- HormonrendszerDocument11 pagesHormonrendszerMíra Petrinovics100% (1)
- LC Kromatográfia - Oktatási SegédanyagDocument7 pagesLC Kromatográfia - Oktatási SegédanyagÉva Nikolett MelegNo ratings yet
- DR. SEFFER ISTVÁN Tudományos MunkásságaDocument24 pagesDR. SEFFER ISTVÁN Tudományos MunkásságaAnna NémethNo ratings yet
- Összes Tétel AnatómiaDocument42 pagesÖsszes Tétel AnatómiaAdrienn KissnéNo ratings yet
- Angina Pectoris Jegyzet MagyarDocument30 pagesAngina Pectoris Jegyzet MagyarRita Hadnagy100% (1)
- A Beöntés CsodájaDocument8 pagesA Beöntés CsodájaultraboxNo ratings yet
- KeringésDocument66 pagesKeringésTóth TamásNo ratings yet
- IzuletekDocument27 pagesIzuletekapi-26149346No ratings yet
- Vérkeringés, Szív És AortaDocument21 pagesVérkeringés, Szív És Aortalilla100% (1)
- Belgy Olah 19 NetDocument62 pagesBelgy Olah 19 NetKoncz AnnaNo ratings yet
- Biokemia BSC 02a SzenhidratokDocument58 pagesBiokemia BSC 02a Szenhidratokanita_nemesNo ratings yet
- Csaki Agnes - Hun - CSONT ES IZULETTAN IZOMTANDocument76 pagesCsaki Agnes - Hun - CSONT ES IZULETTAN IZOMTANKinda LeventeNo ratings yet
- Somogyi Gábor - Igazságügyi ToxikológiaDocument28 pagesSomogyi Gábor - Igazságügyi ToxikológiaDoraNo ratings yet
- Az Elsősegélynyújtás TörténeteDocument8 pagesAz Elsősegélynyújtás TörténeteNikolett KuczóNo ratings yet
- Kutya FizioterapeutaDocument11 pagesKutya FizioterapeutaTesztelek JozsefNo ratings yet
- Biokémia Beugró ÖsszesDocument25 pagesBiokémia Beugró ÖsszesLaura RáczNo ratings yet
- Vér AnatómiájaDocument8 pagesVér AnatómiájaNyisztor RiaNo ratings yet
- 06 Vese BiolDocument116 pages06 Vese BiolAndrea Violetta ÁcsNo ratings yet
- Idegrendszer BetegségeiDocument11 pagesIdegrendszer BetegségeiBrigitta RáczNo ratings yet
- Orvosi Kémia II. 2. Demó AnyagaDocument27 pagesOrvosi Kémia II. 2. Demó Anyagabalage_szNo ratings yet
- 14 Decubitus Felmérés Kezelés Megelőzés ÁpolásDocument9 pages14 Decubitus Felmérés Kezelés Megelőzés ÁpolásRéka BeneNo ratings yet
- 5 1 SzaltelekiDocument85 pages5 1 SzaltelekiRados KatalinNo ratings yet
- Urologia TortenelemDocument28 pagesUrologia TortenelemkereklovagNo ratings yet
- 3.a.hugyuti InfekciokDocument19 pages3.a.hugyuti InfekciokkereklovagNo ratings yet
- UrográfiaDocument2 pagesUrográfiakereklovagNo ratings yet
- P5 Vese BetegségeiDocument87 pagesP5 Vese BetegségeikereklovagNo ratings yet
- Ossz PPT MagyarDocument593 pagesOssz PPT MagyarkereklovagNo ratings yet
- Eloadas 9 MajDocument44 pagesEloadas 9 MajkereklovagNo ratings yet
- Konyv PDFDocument122 pagesKonyv PDFkereklovagNo ratings yet
- 1.fejlodesi Rendellenessegek PDFDocument50 pages1.fejlodesi Rendellenessegek PDFkereklovagNo ratings yet
- Szuleszeti Es Nogyogyaszati PatologiaDocument91 pagesSzuleszeti Es Nogyogyaszati PatologiakereklovagNo ratings yet
- Ultrahang - I. ElőadásDocument14 pagesUltrahang - I. ElőadáskereklovagNo ratings yet
- Eloadas 13 Ovarium PDFDocument15 pagesEloadas 13 Ovarium PDFkereklovagNo ratings yet
- 4 Előadás-CDocument63 pages4 Előadás-CkereklovagNo ratings yet
- Eloadas 12 Noi Nemi SzervekDocument28 pagesEloadas 12 Noi Nemi SzervekkereklovagNo ratings yet
- Eloadas 8 Gyomor-BelekDocument145 pagesEloadas 8 Gyomor-BelekkereklovagNo ratings yet
- Eloadas 7 LegutakDocument106 pagesEloadas 7 LegutakkereklovagNo ratings yet
- Vizkelety o PDFDocument264 pagesVizkelety o PDFkereklovag67% (3)
- MorfopatologiaDocument32 pagesMorfopatologiakereklovagNo ratings yet
- 7 Előadás-28042017cDocument48 pages7 Előadás-28042017ckereklovagNo ratings yet
- 10 Előadás-19052017aDocument80 pages10 Előadás-19052017akereklovagNo ratings yet
- 7 Előadás-28042017cDocument48 pages7 Előadás-28042017ckereklovagNo ratings yet
- ZsiranyagcsereDocument67 pagesZsiranyagcserekereklovagNo ratings yet
- Bara Tivadar Fizikai Vizsgalat Es Sebeszeti Alapfogalmak 1 PDFDocument171 pagesBara Tivadar Fizikai Vizsgalat Es Sebeszeti Alapfogalmak 1 PDFkereklovagNo ratings yet
- 11 Előadas-26052017Document70 pages11 Előadas-26052017kereklovagNo ratings yet
- SejtbioDocument76 pagesSejtbiokereklovagNo ratings yet
- Urologia PDFDocument268 pagesUrologia PDFAnonymous p28Tktgl5No ratings yet
- Sejtbiológia 1Document57 pagesSejtbiológia 1kereklovagNo ratings yet
- AminoglokozidokDocument51 pagesAminoglokozidokkereklovagNo ratings yet
- SedatohipnotikumokDocument24 pagesSedatohipnotikumokkereklovagNo ratings yet
- 1 Eloadas Kardiovaszkularis Anmanezis, TunetekDocument133 pages1 Eloadas Kardiovaszkularis Anmanezis, TunetekkereklovagNo ratings yet