You might also like
- 7 Előadás-28042017cDocument48 pages7 Előadás-28042017ckereklovagNo ratings yet
- Az ImmunrendszerDocument66 pagesAz ImmunrendszerRenáta FerencziNo ratings yet
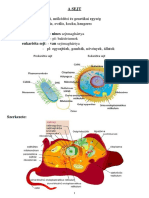
- 09.01.A SejtDocument8 pages09.01.A SejtMolnár GyulaNo ratings yet
- Szív, Erek, VérkörökDocument24 pagesSzív, Erek, VérkörökGombos Angelika Afrodita100% (1)
- Farmakológia Tételek 2Document40 pagesFarmakológia Tételek 2viviNo ratings yet
- LC Kromatográfia - Oktatási SegédanyagDocument7 pagesLC Kromatográfia - Oktatási SegédanyagÉva Nikolett MelegNo ratings yet
- Általános Mikrobiológiai Kérdések KIDOLGOZOTT!Document17 pagesÁltalános Mikrobiológiai Kérdések KIDOLGOZOTT!Bagó IrénNo ratings yet
- Transzfúziós SzabályzatDocument113 pagesTranszfúziós Szabályzatani1995_271012009No ratings yet
- éLETTAN Nyirok Immunolã Gia - 10Document50 pageséLETTAN Nyirok Immunolã Gia - 10KataNo ratings yet
- Vénás VérmintavételDocument3 pagesVénás VérmintavételSzekeres-Csiki KatalinNo ratings yet
- NeurológiaDocument115 pagesNeurológiaKoncz AnnaNo ratings yet
- ABCDE AllapotfelmeresDocument3 pagesABCDE AllapotfelmeresDani Molnar100% (1)
- Edokrin Rendszer...Document32 pagesEdokrin Rendszer...Szabolcs FábiánNo ratings yet
- Emesztorendszer 1Document61 pagesEmesztorendszer 1Ritus91No ratings yet
- Neurológiai Kórképek - 1601129762Document50 pagesNeurológiai Kórképek - 1601129762Monika BurkaNo ratings yet
- Korneth Aniko Klinikai Ismeretek A Belgyogyaszat Koreből Apolonők Reszere - 13Document5 pagesKorneth Aniko Klinikai Ismeretek A Belgyogyaszat Koreből Apolonők Reszere - 13ama001No ratings yet
- 2 Sugárfizikai AlapismeretekDocument31 pages2 Sugárfizikai AlapismeretekHavassy ZoéNo ratings yet
- Vakcinák: Milyen védőoltásokat adassak be gyermekemnek?From EverandVakcinák: Milyen védőoltásokat adassak be gyermekemnek?No ratings yet
- 1.HUN RFRP Script - Vese Fiziológiája, VeseelégtelenségDocument13 pages1.HUN RFRP Script - Vese Fiziológiája, VeseelégtelenségÁkos SzaporánNo ratings yet
- Laborvizsgálati Eredmények Magyarázata KözérthetőenDocument8 pagesLaborvizsgálati Eredmények Magyarázata KözérthetőenCsilla FülöpNo ratings yet
- Dialízis Kezelés GyakorlataDocument116 pagesDialízis Kezelés Gyakorlatatotya91No ratings yet
- A Kivalasztas Szervrendszere Es Endokrinologia WLDocument56 pagesA Kivalasztas Szervrendszere Es Endokrinologia WLNoemi Zold-KemenesNo ratings yet
- Rextra 2014 OnlineDocument148 pagesRextra 2014 OnlineTamásKureczNo ratings yet
- Emésztés Modul IsmertetéseDocument24 pagesEmésztés Modul IsmertetéseBus MelindaNo ratings yet
- Anatómia Tételek - ZsigerekDocument14 pagesAnatómia Tételek - ZsigerekLadiko100% (1)
- Bevarrva: Egy börtönorvos feljegyzései életről és halálrólFrom EverandBevarrva: Egy börtönorvos feljegyzései életről és halálrólNo ratings yet
- A PCOS Gyogyszeres - Kezelesenek - Bizonyos - KerdeseiDocument5 pagesA PCOS Gyogyszeres - Kezelesenek - Bizonyos - KerdeseiAnettNo ratings yet
- Korneth Aniko Klinikai Ismeretek A Belgyogyaszat Koreből Apolonők Reszere - 20Document5 pagesKorneth Aniko Klinikai Ismeretek A Belgyogyaszat Koreből Apolonők Reszere - 20ama001No ratings yet
- 01-Molekularis Biologia TesztDocument64 pages01-Molekularis Biologia TesztMányák-Cseh B. ZitaNo ratings yet
- Bódi Bernadett - Gyakoribb Neurológiai Kórképek Intenzív TerápiájaDocument45 pagesBódi Bernadett - Gyakoribb Neurológiai Kórképek Intenzív Terápiájapeter_soósNo ratings yet
- Tétel (Betegmegfigyelés)Document5 pagesTétel (Betegmegfigyelés)Kiss-Szabados MelindaNo ratings yet
- Kommunikáció A MűtőbenDocument26 pagesKommunikáció A MűtőbenS.AdriNo ratings yet
- Egészségtudományi Alapismeretek II.: 2021/22 Őszi FélévDocument394 pagesEgészségtudományi Alapismeretek II.: 2021/22 Őszi FélévKissné Makai KataNo ratings yet
- Az Idegrendszer BetegségeiDocument79 pagesAz Idegrendszer BetegségeiIzabella TeiglerNo ratings yet
- Biológia Ábrák 1Document21 pagesBiológia Ábrák 1Alexandra NémethNo ratings yet
- Klinikai Farmakológia KVK - LaknerGDocument59 pagesKlinikai Farmakológia KVK - LaknerGKoncz AnnaNo ratings yet
- A Vese ÉlettanaDocument4 pagesA Vese ÉlettanaGesarolNo ratings yet
- TételDocument11 pagesTételZoltánné FarkasNo ratings yet
- Alt KortanDocument31 pagesAlt KortanZoli Künsztler100% (1)
- Alapszakdolgozatok, Mesterszakdolgozatok, Diplomamunkák, Szakdolgozatok, Tudományos Megírása Könnyedén (Tudományos Munka)From EverandAlapszakdolgozatok, Mesterszakdolgozatok, Diplomamunkák, Szakdolgozatok, Tudományos Megírása Könnyedén (Tudományos Munka)No ratings yet
- Egészségügyi AlapismeretekDocument14 pagesEgészségügyi AlapismeretekCordelia CartwrightNo ratings yet
- Tanuljon Hollandul - Gyorsan / Egyszerűen / Hatékonyan: 2000 KulcsszóFrom EverandTanuljon Hollandul - Gyorsan / Egyszerűen / Hatékonyan: 2000 KulcsszóNo ratings yet
- Vér Kórélettana PDFDocument38 pagesVér Kórélettana PDFNikolett NagyNo ratings yet
- XXX. Évfolyam 2022. NyárDocument100 pagesXXX. Évfolyam 2022. NyárVersmondó100% (1)
- Vérkeringés, Szív És AortaDocument21 pagesVérkeringés, Szív És Aortalilla100% (1)
- Regresszív És Progresszív Elváltozások A Sebgyógyulás És Az Oszteoporózis, 5. A Sokk, A Vérzés, A Trombózis És Az EmbolusDocument10 pagesRegresszív És Progresszív Elváltozások A Sebgyógyulás És Az Oszteoporózis, 5. A Sokk, A Vérzés, A Trombózis És Az Embolusdorsolya92No ratings yet
- Extra VasaTioDocument3 pagesExtra VasaTioDani MolnarNo ratings yet
- 8 Előadás-05052017bDocument53 pages8 Előadás-05052017bkereklovagNo ratings yet
- Korneth Aniko Klinikai Ismeretek A Belgyogyaszat Koreből Apolonők Reszere - 15Document5 pagesKorneth Aniko Klinikai Ismeretek A Belgyogyaszat Koreből Apolonők Reszere - 15ama001No ratings yet
- Tanuljon Lengyelül - Gyorsan / Egyszerűen / Hatékonyan: 2000 KulcsszóFrom EverandTanuljon Lengyelül - Gyorsan / Egyszerűen / Hatékonyan: 2000 KulcsszóNo ratings yet
- Emlő Betegségei - 1614981066Document135 pagesEmlő Betegségei - 1614981066Monika BurkaNo ratings yet
- Kiválasztás - TeljesDocument72 pagesKiválasztás - TeljesSarah SimmonsNo ratings yet
- Biokemia BSC 02a SzenhidratokDocument58 pagesBiokemia BSC 02a Szenhidratokanita_nemesNo ratings yet
- Urologia TortenelemDocument28 pagesUrologia TortenelemkereklovagNo ratings yet
- 3.a.hugyuti InfekciokDocument19 pages3.a.hugyuti InfekciokkereklovagNo ratings yet
- UrográfiaDocument2 pagesUrográfiakereklovagNo ratings yet
- P5 Vese BetegségeiDocument87 pagesP5 Vese BetegségeikereklovagNo ratings yet
- Ossz PPT MagyarDocument593 pagesOssz PPT MagyarkereklovagNo ratings yet
- Eloadas 9 MajDocument44 pagesEloadas 9 MajkereklovagNo ratings yet
- Konyv PDFDocument122 pagesKonyv PDFkereklovagNo ratings yet
- 1.fejlodesi Rendellenessegek PDFDocument50 pages1.fejlodesi Rendellenessegek PDFkereklovagNo ratings yet
- Szuleszeti Es Nogyogyaszati PatologiaDocument91 pagesSzuleszeti Es Nogyogyaszati PatologiakereklovagNo ratings yet
- Ultrahang - I. ElőadásDocument14 pagesUltrahang - I. ElőadáskereklovagNo ratings yet
- Eloadas 13 Ovarium PDFDocument15 pagesEloadas 13 Ovarium PDFkereklovagNo ratings yet
- Eloadas 12 Noi Nemi SzervekDocument28 pagesEloadas 12 Noi Nemi SzervekkereklovagNo ratings yet
- Eloadas 8 Gyomor-BelekDocument145 pagesEloadas 8 Gyomor-BelekkereklovagNo ratings yet
- Eloadas 7 LegutakDocument106 pagesEloadas 7 LegutakkereklovagNo ratings yet
- Vizkelety o PDFDocument264 pagesVizkelety o PDFkereklovag67% (3)
- MorfopatologiaDocument32 pagesMorfopatologiakereklovagNo ratings yet
- 8 Előadás-05052017bDocument53 pages8 Előadás-05052017bkereklovagNo ratings yet
- 10 Előadás-19052017aDocument80 pages10 Előadás-19052017akereklovagNo ratings yet
- ZsiranyagcsereDocument67 pagesZsiranyagcserekereklovagNo ratings yet
- Bara Tivadar Fizikai Vizsgalat Es Sebeszeti Alapfogalmak 1 PDFDocument171 pagesBara Tivadar Fizikai Vizsgalat Es Sebeszeti Alapfogalmak 1 PDFkereklovagNo ratings yet
- 11 Előadas-26052017Document70 pages11 Előadas-26052017kereklovagNo ratings yet
- SejtbioDocument76 pagesSejtbiokereklovagNo ratings yet
- Urologia PDFDocument268 pagesUrologia PDFAnonymous p28Tktgl5No ratings yet
- Sejtbiológia 1Document57 pagesSejtbiológia 1kereklovagNo ratings yet
- AminoglokozidokDocument51 pagesAminoglokozidokkereklovagNo ratings yet
- SedatohipnotikumokDocument24 pagesSedatohipnotikumokkereklovagNo ratings yet
- 1 Eloadas Kardiovaszkularis Anmanezis, TunetekDocument133 pages1 Eloadas Kardiovaszkularis Anmanezis, TunetekkereklovagNo ratings yet