You might also like
- Gyakorlas 1Document76 pagesGyakorlas 1Martin Juhász100% (1)
- Probléma Feladatok TémásDocument18 pagesProbléma Feladatok Témásboszoky viragNo ratings yet
- Táplálkozási SzervrendszerDocument20 pagesTáplálkozási Szervrendszergabor23 /No ratings yet
- A Röntgensugár KeletkezéseDocument43 pagesA Röntgensugár KeletkezéseEmese VargaNo ratings yet
- Joseph D'Lacey - HúsDocument177 pagesJoseph D'Lacey - HúsReich Tamara100% (1)
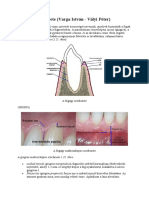
- A Fogágy SzerkezeteDocument8 pagesA Fogágy Szerkezeteterezia99100% (1)
- 11.12. Kidolgozott FeladatokDocument45 pages11.12. Kidolgozott FeladatokbcsmNo ratings yet
- FI-505031202 1 Teljes PDFDocument97 pagesFI-505031202 1 Teljes PDFbcsmNo ratings yet
- Logopedia Tanitasi TervezetekDocument66 pagesLogopedia Tanitasi TervezetekAnikó Joó-Faluvégi100% (4)
- Bevarrva: Egy börtönorvos feljegyzései életről és halálrólFrom EverandBevarrva: Egy börtönorvos feljegyzései életről és halálrólNo ratings yet
- 6 TételDocument2 pages6 TételbcsmNo ratings yet
- Kemia 12 Szerves Reakciok Szaktanari Segedlet Gavlikne Kis AnitaDocument16 pagesKemia 12 Szerves Reakciok Szaktanari Segedlet Gavlikne Kis Anitaszabo anna100% (1)
- Az Allatok Legzese 01Document3 pagesAz Allatok Legzese 01sophieNo ratings yet
- 100 Oras Biologia TematikaDocument3 pages100 Oras Biologia TematikaBors ErzsébetNo ratings yet
- Szerves Kemia AlkinekArenekHeteroaromasHalogenszarm 56Document138 pagesSzerves Kemia AlkinekArenekHeteroaromasHalogenszarm 56Ágnes ZongorNo ratings yet
- Vii - Fogászati Tömőanyagok PDFDocument52 pagesVii - Fogászati Tömőanyagok PDFBattyáni ZsuzsannaNo ratings yet
- Semmelweis Pontszámítás - 2024Document7 pagesSemmelweis Pontszámítás - 2024Zolika0906No ratings yet
- FenymikroszkopiaDocument23 pagesFenymikroszkopiaAugustine CsigiNo ratings yet
- Élettan - 2013 - Hetes JegyzetDocument589 pagesÉlettan - 2013 - Hetes JegyzetRekaNo ratings yet
- ÁllattanDocument28 pagesÁllattanxspatulaxNo ratings yet
- Szaj BetegsegekDocument10 pagesSzaj BetegsegeksplendidflamingoNo ratings yet
- Biofizikia-Kidolgozott-K RD Sek - Szi-F L VDocument23 pagesBiofizikia-Kidolgozott-K RD Sek - Szi-F L VKatalin Strobl100% (1)
- Nukleinsavak BioszferaDocument6 pagesNukleinsavak BioszferaBéla KamuNo ratings yet
- Anatómia 1Document68 pagesAnatómia 1Vilma AddamsNo ratings yet
- A SejtalkotokDocument42 pagesA Sejtalkotokanita_nemesNo ratings yet
- Biokemia II 1Document73 pagesBiokemia II 1Janos GreenMasterNo ratings yet
- Kémia A Mindennapokban - RészösszefoglalásDocument16 pagesKémia A Mindennapokban - RészösszefoglalásKrisztina Vas100% (1)
- A Növények ÉlettanaDocument21 pagesA Növények Élettanakurucz barbaraNo ratings yet
- 2 Het SejtbiologiaDocument42 pages2 Het SejtbiologiaAntal AlexandraNo ratings yet
- FogfehérítésDocument61 pagesFogfehérítésHorvathDavidNo ratings yet
- ORAREND - A Semmelweis Egyetem ÁOK FOK GYTK EKK Karainak OrarendjeDocument2 pagesORAREND - A Semmelweis Egyetem ÁOK FOK GYTK EKK Karainak OrarendjeDrBenjaminButton100% (1)
- Eukarióta És Prokarióta Sejt ÖsszehasonlításaDocument2 pagesEukarióta És Prokarióta Sejt ÖsszehasonlításaFanni Kiss100% (1)
- Altalanos Es Szervetlen KemiaDocument142 pagesAltalanos Es Szervetlen KemiaMárk StrauszNo ratings yet
- Aktív És Passzív Transzport ÖsszehasonlításaDocument2 pagesAktív És Passzív Transzport ÖsszehasonlításaFanni KissNo ratings yet
- Gygyszertan - JegyzetDocument80 pagesGygyszertan - JegyzetcshenszajaNo ratings yet
- KÉMIADocument5 pagesKÉMIAapi-3774250100% (1)
- Nano Kérdések Kidolgozva TeljesDocument23 pagesNano Kérdések Kidolgozva TeljesRebeka VásárhelyiNo ratings yet
- 1.fejezet Alapfogalmak SzámoláshozDocument53 pages1.fejezet Alapfogalmak SzámoláshozVargaingrid100% (1)
- Emelt Kémia Szóbeli Altétel (C) Ötletek Nem Kidolgozott - HonlapraDocument11 pagesEmelt Kémia Szóbeli Altétel (C) Ötletek Nem Kidolgozott - Honlaprafelkészítés KemBioNo ratings yet
- Kémia FogalomtárDocument26 pagesKémia FogalomtárErica Gy VargaNo ratings yet
- A Kiválasztó Szervrendszer Anatómiája, A Kiválasztás Élettana.Document44 pagesA Kiválasztó Szervrendszer Anatómiája, A Kiválasztás Élettana.Szekeres-Kovács PetraNo ratings yet
- Nem Szteroid GyulladáscsökkentőkDocument28 pagesNem Szteroid GyulladáscsökkentőkRoland JuonNo ratings yet
- Szervetlen Jegyzet - KepekkelDocument414 pagesSzervetlen Jegyzet - KepekkelSzabó Tibor75% (4)
- Betegmegfigy Es Vit Param - ágazATI VIZSGADocument64 pagesBetegmegfigy Es Vit Param - ágazATI VIZSGAKutasi PiroskaNo ratings yet
- Folyadékok És GázokDocument18 pagesFolyadékok És Gázokapi-3828155100% (5)
- Biologia Onellenőrző Feladatok Megoldasokkal PDFDocument40 pagesBiologia Onellenőrző Feladatok Megoldasokkal PDFZsagikaNo ratings yet
- Kemia 11 Emelt Szintu Kiserletek Szaktanari Zseni ZsofiaDocument16 pagesKemia 11 Emelt Szintu Kiserletek Szaktanari Zseni Zsofiaszabo anna100% (1)
- SzervesII 2018 2019 PDFDocument275 pagesSzervesII 2018 2019 PDFHelgert Bettina ÁgnesNo ratings yet
- Kémia MinimumDocument28 pagesKémia MinimumLídia AnikNo ratings yet
- BetalaktamokDocument34 pagesBetalaktamokkereklovagNo ratings yet
- Fizika Fogalomtar 2011-05-17Document315 pagesFizika Fogalomtar 2011-05-17Tímea TokodiNo ratings yet
- ElektrokémiaDocument175 pagesElektrokémiaFanni MegyeriNo ratings yet
- 11 PopulaciokkolcsonhatasaiDocument13 pages11 PopulaciokkolcsonhatasaiEmília NagyNo ratings yet
- Biofizika TételekDocument75 pagesBiofizika TételekVas Lilla100% (1)
- Biokemia BSC 01 EnzimekDocument48 pagesBiokemia BSC 01 Enzimekanita_nemesNo ratings yet
- 2.2 A Sejtek Anyagcsere FolyamataiDocument8 pages2.2 A Sejtek Anyagcsere FolyamataiRichard CortezNo ratings yet
- Modern Fizika LaboratóriumDocument276 pagesModern Fizika LaboratóriumBöhm úrNo ratings yet
- Érettségire EgyenletekDocument126 pagesÉrettségire Egyenletekanita_nemesNo ratings yet
- Növényi SzövetekDocument3 pagesNövényi SzövetekGombos Angelika AfroditaNo ratings yet
- Harasztok AlapDocument4 pagesHarasztok AlapBors ErzsébetNo ratings yet
- Kemiai ReakciokDocument42 pagesKemiai ReakciokAdorján Vas100% (1)
- Idegrendszer 1. RészDocument30 pagesIdegrendszer 1. RészbcsmNo ratings yet
- NyelvtanDocument2 pagesNyelvtanbcsmNo ratings yet
- FizikapéldatárDocument230 pagesFizikapéldatárHalász Gergely100% (2)
- FizikapéldatárDocument230 pagesFizikapéldatárHalász Gergely100% (2)
- HalakDocument2 pagesHalakbcsmNo ratings yet
- Netes SzakácskönyvDocument127 pagesNetes SzakácskönyvbcsmNo ratings yet