
You might also like
- Solid Water Particles as Building Blocks of Meridians: 構建經絡的固態水粒子From EverandSolid Water Particles as Building Blocks of Meridians: 構建經絡的固態水粒子No ratings yet
- 1999 Paper 2 MarkingDocument11 pages1999 Paper 2 Markingmichelle005410No ratings yet
- 水生 水耕Document11 pages水生 水耕Zi-Yu HongNo ratings yet
- 1 1 1 8 2021生物中考复习手册Document30 pages1 1 1 8 2021生物中考复习手册Lia CNo ratings yet
- 植物的营养与运输大纲Document5 pages植物的营养与运输大纲Henry LiNo ratings yet
- 925808 國中自然第一冊 學習手冊 PDFDocument69 pages925808 國中自然第一冊 學習手冊 PDFKenNo ratings yet
- 國中自然第一冊 學習手冊Document69 pages國中自然第一冊 學習手冊goodman.ks67% (3)
- 0 soil 土壤棲地Document16 pages0 soil 土壤棲地林雨莊No ratings yet
- 植物生理学Document42 pages植物生理学AndelingNo ratings yet
- UntitledDocument28 pagesUntitledHuajidawang 666No ratings yet
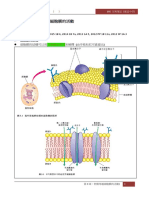
- Cell Membrane: Daniel ZhuDocument19 pagesCell Membrane: Daniel ZhuDaniel ZhuNo ratings yet
- 微生物学 (沈萍) 考试重点Document42 pages微生物学 (沈萍) 考试重点qiyuding58No ratings yet
- 1992 Marking Paper 2Document9 pages1992 Marking Paper 2michelle005410No ratings yet
- 生物Document95 pages生物foongNo ratings yet
- 单元四 植物Document31 pages单元四 植物Puie KhongNo ratings yet
- 生物小练习册错题Document3 pages生物小练习册错题KORENNo ratings yet
- 习题Document120 pages习题anon-580967No ratings yet
- 會考生物筆記Document7 pages會考生物筆記Jenny ChoiNo ratings yet
- 全植物生理Document8 pages全植物生理ZhiNo ratings yet
- 111中医汗法现代生理病理基础及临床意义探讨 肖党生Document3 pages111中医汗法现代生理病理基础及临床意义探讨 肖党生banbanroaNo ratings yet
- 四年级科学笔记 KSSR SemakanDocument4 pages四年级科学笔记 KSSR SemakanGAN QI LI MoeNo ratings yet
- 03 物質穿越細胞膜的活動 2021docxDocument16 pages03 物質穿越細胞膜的活動 2021docx归次100% (1)
- 生物 选修二 知识点总结Document21 pages生物 选修二 知识点总结a1280424159No ratings yet
- 生物大书表格Document3 pages生物大书表格KORENNo ratings yet
- 归纳3+细胞怎样构成生物体 【必背知识】2021 2022学年七年级生物上册知识点归纳(人教版)Document5 pages归纳3+细胞怎样构成生物体 【必背知识】2021 2022学年七年级生物上册知识点归纳(人教版)Han DuNo ratings yet
- 高三生物1Document15 pages高三生物1Lizzy YuNo ratings yet
- 4 - 细胞Document76 pages4 - 细胞Davino Alcantara Dores NetoNo ratings yet
- 5P14植物營養學Document13 pages5P14植物營養學T4醫學護理類編輯室No ratings yet
- 2月Class 初一 科学5 Learning Material1Document2 pages2月Class 初一 科学5 Learning Material1MintzNo ratings yet
- Ch02植物體的形態、構造與功能 課本pdf檔Document46 pagesCh02植物體的形態、構造與功能 課本pdf檔洪允升No ratings yet
- 期末中文翻譯Document8 pages期末中文翻譯s110065011No ratings yet
- KSSR 科学笔记Document35 pagesKSSR 科学笔记Chua Geok HOONNo ratings yet
- 土壤 活的有機體Document23 pages土壤 活的有機體chientai0% (1)
- 2-1 Point (105) A0Document13 pages2-1 Point (105) A0b104093038 (TMU)No ratings yet
- 人体解剖生理学 (第三版)Document261 pages人体解剖生理学 (第三版)Auther Merlin100% (1)
- SC f1 (Ac)Document48 pagesSC f1 (Ac)Tee Qi ENNo ratings yet
- ch03c BioDocument8 pagesch03c BioGENIUS GGECNo ratings yet
- 光合作用 1Document26 pages光合作用 1tehpNo ratings yet
- 2022教科版五年级下册科学知识点总结与归纳Document8 pages2022教科版五年级下册科学知识点总结与归纳AliceGaoNo ratings yet
- 第4章 真核生物Document141 pages第4章 真核生物评论员No ratings yet
- Bio Jr Ppt - Plant Respiration 植物呼吸 2022Document96 pagesBio Jr Ppt - Plant Respiration 植物呼吸 2022aman97881234No ratings yet
- 单元4 植物Document51 pages单元4 植物KOAY SHU CHING MoeNo ratings yet
- 2017科学重点笔记Document17 pages2017科学重点笔记高兴No ratings yet
- 2017科学重点笔记 PDFDocument17 pages2017科学重点笔记 PDF高兴100% (1)
- UPSR SC NotesDocument17 pagesUPSR SC NotesH'NG EE JOE MoeNo ratings yet
- 初一科学 第六章 复习Document9 pages初一科学 第六章 复习S7〆 jerryNo ratings yet
- Score ADocument91 pagesScore ARin Chorungreang RinNo ratings yet
- 4 第4章细胞质膜 翟中和第四版 65Document65 pages4 第4章细胞质膜 翟中和第四版 65xiongliangyao1992No ratings yet
- 细胞的结构与功能中文版综合笔记Document8 pages细胞的结构与功能中文版综合笔记高二理1(38)李祥俊LEE XIANG JUNNo ratings yet
- 第三章講義 CH03 - 72K PDFDocument72 pages第三章講義 CH03 - 72K PDF廖敏伊No ratings yet
- 【報告】1998 馬志珍 海產餌用微藻固定化保種技術Document5 pages【報告】1998 馬志珍 海產餌用微藻固定化保種技術realgigiNo ratings yet
- Pi Water Structured WaterDocument8 pagesPi Water Structured WaterLetsdoit EczemaNo ratings yet
- KXDocument6 pagesKX锺欣锜No ratings yet
- Inhibition Effect of Aquaculture Water of Salvinia Natans (L ) All On Microcystis Aeruginosa PCC7806Document6 pagesInhibition Effect of Aquaculture Water of Salvinia Natans (L ) All On Microcystis Aeruginosa PCC7806LouiseNo ratings yet
- 第2章 原核微生物Document104 pages第2章 原核微生物评论员No ratings yet
- 生命科学研究中常用模式生物 PDFDocument10 pages生命科学研究中常用模式生物 PDFHaowen WuNo ratings yet
- 鳞翅目昆虫肠道微生物的多样性及其与宿主的相互作用Document13 pages鳞翅目昆虫肠道微生物的多样性及其与宿主的相互作用tabaci BemisiaNo ratings yet
- nphssf2014-080311Document32 pagesnphssf2014-080311Chien Hung YenNo ratings yet
- 妈妈出的科学?Document3 pages妈妈出的科学?m-13172411No ratings yet
- Chapter 3 Biological UnitDocument37 pagesChapter 3 Biological UnitMian Yu LimNo ratings yet
- Once Upon a Time... (Chinesische Ausgabe): a Scientific Fairy Tale - Volume I - (Chinesisch)From EverandOnce Upon a Time... (Chinesische Ausgabe): a Scientific Fairy Tale - Volume I - (Chinesisch)No ratings yet
- Yin Yang and Wuxing (Five Elements) and System Control Theory: 宏观物理学基本原理From EverandYin Yang and Wuxing (Five Elements) and System Control Theory: 宏观物理学基本原理No ratings yet
- Dr. Jizhou Kang's Information Medicine - The Handbook: A 60 year experience of Organic Integration of Chinese and Western Medicine (Volume 2): 康氏信息医学──中医学西医学三融合(下册)From EverandDr. Jizhou Kang's Information Medicine - The Handbook: A 60 year experience of Organic Integration of Chinese and Western Medicine (Volume 2): 康氏信息医学──中医学西医学三融合(下册)No ratings yet
- Alternative Cancer Treatment for 21th Century - The Untold Truth About Cancer: 二十一世纪的抗癌新法 ─ 醫生不會告訴你的癌症真相From EverandAlternative Cancer Treatment for 21th Century - The Untold Truth About Cancer: 二十一世纪的抗癌新法 ─ 醫生不會告訴你的癌症真相No ratings yet
- ISOQOL生存质量与健康结局测量词典: 简体中文翻译本 第一版From EverandISOQOL生存质量与健康结局测量词典: 简体中文翻译本 第一版Nancy MayoNo ratings yet
- Constructive Realism in Chinese MedicineFrom EverandConstructive Realism in Chinese MedicineFriedrich G. WallnerNo ratings yet
- An Interpretation of Gravitational Anomaly Using New Formulae On Gravity And Repulsion: 新引力斥力公式对引力异常现象的解读From EverandAn Interpretation of Gravitational Anomaly Using New Formulae On Gravity And Repulsion: 新引力斥力公式对引力异常现象的解读No ratings yet



















































































![新興建築技術 [Chinese (Traditional)]](https://imgv2-1-f.scribdassets.com/img/series/539156997/149x198/3c801d2cf3/1677532549?v=1)

