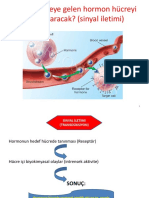
You might also like
- Steroid Hormonlar Ve ReseptorleriDocument41 pagesSteroid Hormonlar Ve ReseptorleriRugeyye CanNo ratings yet
- Hormonların Genel Özellikleri: Prof. Dr. Hafize UZUN Tıbbi Biyokimya ADDocument48 pagesHormonların Genel Özellikleri: Prof. Dr. Hafize UZUN Tıbbi Biyokimya AD230101026No ratings yet
- 1) Endokrin Sistem Farmakolojisine Giriş.zsDocument69 pages1) Endokrin Sistem Farmakolojisine Giriş.zshakan_b91No ratings yet
- Hücrei̇çi̇ Si̇nyal İleti̇m Si̇stemleri̇Document91 pagesHücrei̇çi̇ Si̇nyal İleti̇m Si̇stemleri̇CevikAlp93% (14)
- Hucreler Arasi Sinyal Iletim Mekanizmalari YeniDocument42 pagesHucreler Arasi Sinyal Iletim Mekanizmalari YeniMine İpekNo ratings yet
- 2 Sinyal Iletimi 2020 TÃ RKDocument77 pages2 Sinyal Iletimi 2020 TÃ RKheroma heromaNo ratings yet
- Hücrede Si̇nyal İleti̇mi̇Document75 pagesHücrede Si̇nyal İleti̇mi̇Ibrahim DurmuşNo ratings yet
- 5.sinyal IletimiDocument31 pages5.sinyal Iletimieren badurNo ratings yet
- Santral Sinir Sistemi FarmakolojisiDocument287 pagesSantral Sinir Sistemi FarmakolojisiSTETUSKOP.COM-TIP VE TUS'UN MERKEZİ100% (1)
- HAFTA - Hücresel Sinyal IletimiDocument33 pagesHAFTA - Hücresel Sinyal Iletimizy.shitsiteNo ratings yet
- Hücrelerin Kimyasal Habercilerle KontrolüDocument27 pagesHücrelerin Kimyasal Habercilerle KontrolüRecep UçakNo ratings yet
- İkincil Haberciler SonDocument62 pagesİkincil Haberciler Sonfatmadinc7777No ratings yet
- BIYOKIMYADocument68 pagesBIYOKIMYAEsra OzdemirNo ratings yet
- BİY448 Konu-10Document17 pagesBİY448 Konu-10eNo ratings yet
- Pharmacodynamics 080422Document23 pagesPharmacodynamics 080422Merve kdkkNo ratings yet
- İlaçlarin Etki Mekanizmalari: Prof. Dr. Ayşe EROL Ege Üniversitesi Tıp Fakültesi Tıbbi Farmakoloji Anabilim DalıDocument21 pagesİlaçlarin Etki Mekanizmalari: Prof. Dr. Ayşe EROL Ege Üniversitesi Tıp Fakültesi Tıbbi Farmakoloji Anabilim DalıAyça ÇakılNo ratings yet
- Hücre Biyolojisi 2 / Sinyal İletimi 1Document34 pagesHücre Biyolojisi 2 / Sinyal İletimi 1zuhalNo ratings yet
- Sinyal Ileti Yollari-3 - TGFbetaDocument30 pagesSinyal Ileti Yollari-3 - TGFbetamaestro1234567890No ratings yet
- Gen Ifadesinin Düzenlenmesi Dr. Behice ZERENDocument78 pagesGen Ifadesinin Düzenlenmesi Dr. Behice ZERENbehice zerenNo ratings yet
- Altun FarmakolojiDocument400 pagesAltun FarmakolojiElmira ShamilovaNo ratings yet
- Operon HipoteziDocument33 pagesOperon HipoteziZeynep Günay100% (1)
- HormonlarDocument51 pagesHormonlarScafNo ratings yet
- 8 EnzimlerDocument70 pages8 EnzimleraguzelenkaNo ratings yet
- Kortikosteroidler Ve Antagonistleri: Prof. Dr. Tijen Kaya TEMİZDocument42 pagesKortikosteroidler Ve Antagonistleri: Prof. Dr. Tijen Kaya TEMİZAhmet OkulNo ratings yet
- Santral Sinir Sistemi Farmakolojisinin Temelleri: Doç - Dr.M.Kemal YıldırımDocument59 pagesSantral Sinir Sistemi Farmakolojisinin Temelleri: Doç - Dr.M.Kemal YıldırımALTAN26No ratings yet
- 1.D II Hormon Genel ÖzellikDocument56 pages1.D II Hormon Genel ÖzellikkaanNo ratings yet
- Ökaryotlarda Transkripsiyonel Düzenlenmenin Prensipleri Dr. Behice ZERENDocument45 pagesÖkaryotlarda Transkripsiyonel Düzenlenmenin Prensipleri Dr. Behice ZERENbehice zerenNo ratings yet
- Prokaryotlarda Transkripsiyonel Düzenlenmenin Prensipleri Dr. Behice ZERENDocument44 pagesProkaryotlarda Transkripsiyonel Düzenlenmenin Prensipleri Dr. Behice ZERENbehice zerenNo ratings yet
- Geneti̇k Bi̇lgi̇ AkişiDocument119 pagesGeneti̇k Bi̇lgi̇ Akişikmdv1No ratings yet
- 13 5HTDocument52 pages13 5HTBarış Mahir GültekinNo ratings yet
- Modül 13 Birleşmiş 2Document456 pagesModül 13 Birleşmiş 2bengNo ratings yet
- 9 HormonlarDocument188 pages9 HormonlarZeynep GünayNo ratings yet
- Genel CerrahiDocument206 pagesGenel Cerrahioncetus100% (1)
- Hafta Hücrede Sinyal İletimiDocument124 pagesHafta Hücrede Sinyal İletimisuedacoban18No ratings yet
- Okaryotlarda Transkripsiyon Kontrolu - Muk - 12.02.2021Document45 pagesOkaryotlarda Transkripsiyon Kontrolu - Muk - 12.02.2021Serenay ParlakNo ratings yet
- BIK 158-22 EnzimlerDocument14 pagesBIK 158-22 EnzimlerMona Ghasem zadeNo ratings yet
- HormonlarDocument115 pagesHormonlarAsraNo ratings yet
- Farmakoloji Terimleri SözlüğüDocument44 pagesFarmakoloji Terimleri Sözlüğüarathorn5100% (1)
- Sinyal İletimi2Document26 pagesSinyal İletimi2Yagmur KirazNo ratings yet
- ENZİMLERDocument26 pagesENZİMLERezgigeyik02No ratings yet
- Hücrede Protein Sentezi Ve PTMDocument31 pagesHücrede Protein Sentezi Ve PTMErdem DersNo ratings yet
- Enzimler 1Document54 pagesEnzimler 1براء الشهابيNo ratings yet
- Hücreler Arasi Haberleşme Ve İleti̇şi̇mDocument34 pagesHücreler Arasi Haberleşme Ve İleti̇şi̇mTolga Zorlu75% (4)
- Prokaryotlarda Gen Ifadesinin Duzenlenmesi - Muk-10.02.2021Document69 pagesProkaryotlarda Gen Ifadesinin Duzenlenmesi - Muk-10.02.2021Serenay ParlakNo ratings yet
- Hücreler Arası IletişimDocument37 pagesHücreler Arası IletişimSoydan DoğanNo ratings yet
- Pro Gen 10Document79 pagesPro Gen 10Enes BozdağNo ratings yet
- Gen İfadesi̇ni̇n KontrolüDocument23 pagesGen İfadesi̇ni̇n Kontrolütibbigenetik67No ratings yet
- 06 - Pankreas Ve GİS Hormonları 2Document20 pages06 - Pankreas Ve GİS Hormonları 2Recep Taha ErdoganNo ratings yet
- 2018 Endokri̇n Si̇stem Fi̇zyoloji̇si̇Document25 pages2018 Endokri̇n Si̇stem Fi̇zyoloji̇si̇Sean KirkNo ratings yet
- Enzimatik Reaksiyon ÖrnekleriDocument53 pagesEnzimatik Reaksiyon ÖrnekleriCeyhun YücelNo ratings yet
- 19-Hormon MetabolizmasıDocument54 pages19-Hormon MetabolizmasıErhan KarabulutNo ratings yet
- Gen Ve Genom YapisiDocument5 pagesGen Ve Genom Yapisicigdem1985No ratings yet
- Sizofreni Tedavisinde Güncel GelişmelerDocument17 pagesSizofreni Tedavisinde Güncel GelişmelerEmrah Songur100% (2)
- 10-Translasyon Ve Post Translayonel ModifikasyonlarDocument38 pages10-Translasyon Ve Post Translayonel Modifikasyonlargdhvf fbjfghNo ratings yet
- Sitokrom P450 Enzim Sistemi Ve İlaç - 20131204095139Document15 pagesSitokrom P450 Enzim Sistemi Ve İlaç - 20131204095139Shifa Minhatun Niza'No ratings yet
- Genomik-Ökaryot GenomuDocument39 pagesGenomik-Ökaryot Genomuaminebilge20003No ratings yet