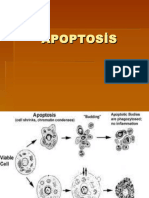
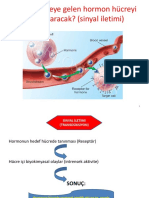
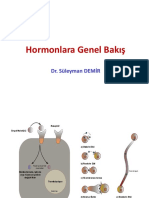
You might also like
- Ökaryotlarda Transkripsiyonel Düzenlenmenin Prensipleri Dr. Behice ZERENDocument45 pagesÖkaryotlarda Transkripsiyonel Düzenlenmenin Prensipleri Dr. Behice ZERENbehice zerenNo ratings yet
- Gen İfadesi̇ni̇n KontrolüDocument23 pagesGen İfadesi̇ni̇n Kontrolütibbigenetik67No ratings yet
- Öka Gen 11Document123 pagesÖka Gen 11Enes BozdağNo ratings yet
- Operon HipoteziDocument33 pagesOperon HipoteziZeynep Günay100% (1)
- Pro Gen 10Document79 pagesPro Gen 10Enes BozdağNo ratings yet
- Okaryotlarda Transkripsiyon Kontrolu - Muk - 12.02.2021Document45 pagesOkaryotlarda Transkripsiyon Kontrolu - Muk - 12.02.2021Serenay ParlakNo ratings yet
- Hücrede Si̇nyal İleti̇mi̇Document75 pagesHücrede Si̇nyal İleti̇mi̇Ibrahim DurmuşNo ratings yet
- Gen Ve Genom YapisiDocument5 pagesGen Ve Genom Yapisicigdem1985No ratings yet
- Gen Ifadesinin Düzenlenmesi Dr. Behice ZERENDocument78 pagesGen Ifadesinin Düzenlenmesi Dr. Behice ZERENbehice zerenNo ratings yet
- Prokaryotlarda Gen Ifadesinin Duzenlenmesi - Muk-10.02.2021Document69 pagesProkaryotlarda Gen Ifadesinin Duzenlenmesi - Muk-10.02.2021Serenay ParlakNo ratings yet
- Sinyal İletimi2Document26 pagesSinyal İletimi2Yagmur KirazNo ratings yet
- 9-11.hafta Populasyon Genetigi Ders NotlarıDocument30 pages9-11.hafta Populasyon Genetigi Ders Notlarıintizarguzelcay06No ratings yet
- Hücre Biyolojisi 2 / Sinyal İletimi 1Document34 pagesHücre Biyolojisi 2 / Sinyal İletimi 1zuhalNo ratings yet
- Hafta 11Document21 pagesHafta 11Fatma İrem AtaolNo ratings yet
- Hücreler Arası IletişimDocument37 pagesHücreler Arası IletişimSoydan DoğanNo ratings yet
- Steroid Hormonlar Ve ReseptorleriDocument41 pagesSteroid Hormonlar Ve ReseptorleriRugeyye CanNo ratings yet
- Kanser BiyokimyasıDocument90 pagesKanser BiyokimyasıMurtaza KurtadamNo ratings yet
- Hafta4 Sinir Sistemi 2Document25 pagesHafta4 Sinir Sistemi 2r6ctdw22npNo ratings yet
- 6 - Hücre Ölümsüzlüğü Ve TümörleşmeDocument74 pages6 - Hücre Ölümsüzlüğü Ve TümörleşmeFurkan YalandanNo ratings yet
- Onkogenler Tumor Supressor GenlerDocument72 pagesOnkogenler Tumor Supressor GenlerNadir KoçakNo ratings yet
- Hücrede Protein Sentezi Ve PTMDocument31 pagesHücrede Protein Sentezi Ve PTMErdem DersNo ratings yet
- Hücreler Arasi Haberleşme Ve İleti̇şi̇mDocument34 pagesHücreler Arasi Haberleşme Ve İleti̇şi̇mTolga Zorlu75% (4)
- Hormon Etki Mekanizmaları: Prof. Dr. Hafize UZUN Tıbbi Biyokimya ADDocument59 pagesHormon Etki Mekanizmaları: Prof. Dr. Hafize UZUN Tıbbi Biyokimya AD230101026No ratings yet
- Histoloji Ve Embiryoloji SunumDocument31 pagesHistoloji Ve Embiryoloji Sunumpolatseda159No ratings yet
- 1 Hücre Döngüsü Ve Kontrol Noktaları SonDocument44 pages1 Hücre Döngüsü Ve Kontrol Noktaları SonemirxqqNo ratings yet
- Hücrelerde SinyalDocument24 pagesHücrelerde SinyalmustafasacarNo ratings yet
- Tek Zarla Kapli OrganellerDocument130 pagesTek Zarla Kapli Organeller2bv6yyhwtzNo ratings yet
- İkincil Haberciler SonDocument62 pagesİkincil Haberciler Sonfatmadinc7777No ratings yet
- Hormonların Genel Özellikleri: Prof. Dr. Hafize UZUN Tıbbi Biyokimya ADDocument48 pagesHormonların Genel Özellikleri: Prof. Dr. Hafize UZUN Tıbbi Biyokimya AD230101026No ratings yet
- Hucreler Arasi Sinyal Iletim Mekanizmalari YeniDocument42 pagesHucreler Arasi Sinyal Iletim Mekanizmalari YeniMine İpekNo ratings yet
- Prokaryotlarda Transkripsiyonel Düzenlenmenin Prensipleri Dr. Behice ZERENDocument44 pagesProkaryotlarda Transkripsiyonel Düzenlenmenin Prensipleri Dr. Behice ZERENbehice zerenNo ratings yet
- Hucre Kulturunde Transfeksiyon TeknikleriDocument3 pagesHucre Kulturunde Transfeksiyon Teknikleribio_engineer50% (4)
- 13-Genom VaryasyonlariDocument56 pages13-Genom Varyasyonlarigdhvf fbjfghNo ratings yet
- Hafta Hücrede Sinyal İletimiDocument124 pagesHafta Hücrede Sinyal İletimisuedacoban18No ratings yet
- 10-Translasyon Ve Post Translayonel ModifikasyonlarDocument38 pages10-Translasyon Ve Post Translayonel Modifikasyonlargdhvf fbjfghNo ratings yet
- Immunogenetik / 20.05.2011Document56 pagesImmunogenetik / 20.05.2011zuhalNo ratings yet
- 1.D II Hormon Genel ÖzellikDocument56 pages1.D II Hormon Genel ÖzellikkaanNo ratings yet
- UntitledDocument23 pagesUntitledBeyzanur TümerNo ratings yet
- Endokrin SistemDocument41 pagesEndokrin SistemNisa KutlukkayaNo ratings yet
- Hücreler Arasi İleti̇şi̇mDocument39 pagesHücreler Arasi İleti̇şi̇mvolki777100% (2)
- Histoloji Fizyoloji EmbriyolojiDocument255 pagesHistoloji Fizyoloji Embriyolojioncetus100% (1)
- Prokaryotlarda Gen İfadesi̇ni̇n Düzenlenmesi̇Document15 pagesProkaryotlarda Gen İfadesi̇ni̇n Düzenlenmesi̇biyoprosesNo ratings yet
- Mutasyon 18Document59 pagesMutasyon 18Enes BozdağNo ratings yet
- Konu 7Document65 pagesKonu 7metehan canseverNo ratings yet
- Preteomik FinalDocument6 pagesPreteomik Finalgptbc9xmnhNo ratings yet
- Hücrei̇çi̇ Si̇nyal İleti̇m Si̇stemleri̇Document91 pagesHücrei̇çi̇ Si̇nyal İleti̇m Si̇stemleri̇CevikAlp93% (14)
- Transkripsiyon VE TranslasyonDocument39 pagesTranskripsiyon VE Translasyonenes özNo ratings yet
- Mikoloji 5Document31 pagesMikoloji 526ugurNo ratings yet
- 5-TOKSİKOGENOMİK Ve GENETİK NEDENLERE BAĞLI TOKSİSİTEDocument120 pages5-TOKSİKOGENOMİK Ve GENETİK NEDENLERE BAĞLI TOKSİSİTEŞerif AliNo ratings yet
- 5.sinyal IletimiDocument31 pages5.sinyal Iletimieren badurNo ratings yet
- UE-YL Iş Hijyen FRK Ders 8Document30 pagesUE-YL Iş Hijyen FRK Ders 8Cağdaş AydınNo ratings yet
- Epigenetik Modifikasyonlar Ve Erkek InfertilitesiDocument68 pagesEpigenetik Modifikasyonlar Ve Erkek InfertilitesitosssoNo ratings yet
- Modül 13 Birleşmiş 2Document456 pagesModül 13 Birleşmiş 2bengNo ratings yet
- Hücrelerin Kimyasal Habercilerle KontrolüDocument27 pagesHücrelerin Kimyasal Habercilerle KontrolüRecep UçakNo ratings yet
- Hücre DöngüsüDocument132 pagesHücre Döngüsüzuhal100% (2)
- Yardımcı Üreme Teknikleri Seçmeli Dersi - DMRDocument35 pagesYardımcı Üreme Teknikleri Seçmeli Dersi - DMRŞahin GündoğduNo ratings yet