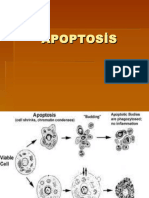
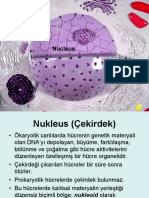
You might also like
- Öka Gen 11Document123 pagesÖka Gen 11Enes BozdağNo ratings yet
- Prokaryotlarda Gen Ifadesinin Duzenlenmesi - Muk-10.02.2021Document69 pagesProkaryotlarda Gen Ifadesinin Duzenlenmesi - Muk-10.02.2021Serenay ParlakNo ratings yet
- 8 TranskripsiyonDocument75 pages8 Transkripsiyongdhvf fbjfghNo ratings yet
- Ökaryotlarda Transkripsiyonel Düzenlenmenin Prensipleri Dr. Behice ZERENDocument45 pagesÖkaryotlarda Transkripsiyonel Düzenlenmenin Prensipleri Dr. Behice ZERENbehice zerenNo ratings yet
- Gen Ve Genom YapisiDocument5 pagesGen Ve Genom Yapisicigdem1985No ratings yet
- Preteomik FinalDocument6 pagesPreteomik Finalgptbc9xmnhNo ratings yet
- Gen Anlatımı 1 TranskripsiyonDocument71 pagesGen Anlatımı 1 Transkripsiyonzuhal100% (3)
- Prokaryotlarda Transkripsiyonel Düzenlenmenin Prensipleri Dr. Behice ZERENDocument44 pagesProkaryotlarda Transkripsiyonel Düzenlenmenin Prensipleri Dr. Behice ZERENbehice zerenNo ratings yet
- 0809 1 4 07GenIfadesiDocument68 pages0809 1 4 07GenIfadesiAyşe Aksu ÖzcanNo ratings yet
- Geneti̇k Bi̇lgi̇ AkişiDocument119 pagesGeneti̇k Bi̇lgi̇ Akişikmdv1No ratings yet
- Bakteri GenetiğiDocument70 pagesBakteri Genetiğiمحمد محمد حتحوتNo ratings yet
- 9 - Transkripsiyon TranslasyonDocument36 pages9 - Transkripsiyon TranslasyonmerveNo ratings yet
- Transkripsiyon Sonrası ModifikasyonlarDocument22 pagesTranskripsiyon Sonrası ModifikasyonlarÖmer LaçinNo ratings yet
- 9-11.hafta Populasyon Genetigi Ders NotlarıDocument30 pages9-11.hafta Populasyon Genetigi Ders Notlarıintizarguzelcay06No ratings yet
- Okaryotlarda Transkripsiyon Kontrolu - Muk - 12.02.2021Document45 pagesOkaryotlarda Transkripsiyon Kontrolu - Muk - 12.02.2021Serenay ParlakNo ratings yet
- Gen Ifadesinin Düzenlenmesi Dr. Behice ZERENDocument78 pagesGen Ifadesinin Düzenlenmesi Dr. Behice ZERENbehice zerenNo ratings yet
- TranskripsiyonDocument39 pagesTranskripsiyonDamla KuruNo ratings yet
- Genomik-Ökaryot GenomuDocument39 pagesGenomik-Ökaryot Genomuaminebilge20003No ratings yet
- Moleküler Biyoloji H7sDocument57 pagesMoleküler Biyoloji H7sr67cm777vwNo ratings yet
- Operon HipoteziDocument33 pagesOperon HipoteziZeynep Günay100% (1)
- 12-Genetik Sifre Ve Kontrol MekanizmalariDocument69 pages12-Genetik Sifre Ve Kontrol Mekanizmalarigdhvf fbjfghNo ratings yet
- Transkripsiyon VE TranslasyonDocument39 pagesTranskripsiyon VE Translasyonenes özNo ratings yet
- Hücrede Protein Sentezi Ve PTMDocument31 pagesHücrede Protein Sentezi Ve PTMErdem DersNo ratings yet
- MEA'Ya SaygılarımlaDocument3 pagesMEA'Ya Saygılarımlahiranurkaygusuz13No ratings yet
- Dna Repli̇kasyonuDocument62 pagesDna Repli̇kasyonuCevikAlpNo ratings yet
- Ders (Tıbbi Biyoloji)Document39 pagesDers (Tıbbi Biyoloji)rzcxhbpbq2No ratings yet
- Nükleik AsitlerDocument32 pagesNükleik Asitlerbilaltorpa49No ratings yet
- Tinywow - 4) 2022 Mitokondri Ve Peroksizom - 38179544Document46 pagesTinywow - 4) 2022 Mitokondri Ve Peroksizom - 38179544Rodoric Koumato (Lerodo)No ratings yet
- Prokaryot, Ökaryot Ve Virüs Dr. Behice ZERENDocument44 pagesProkaryot, Ökaryot Ve Virüs Dr. Behice ZERENbehice zerenNo ratings yet
- Immunogenetik / 20.05.2011Document56 pagesImmunogenetik / 20.05.2011zuhalNo ratings yet
- Mikoloji 5Document31 pagesMikoloji 526ugurNo ratings yet
- Kodlanmayan RnaDocument15 pagesKodlanmayan RnaAleyna Göknil GÖKSUNo ratings yet
- Gen İfadesi̇ni̇n KontrolüDocument23 pagesGen İfadesi̇ni̇n Kontrolütibbigenetik67No ratings yet
- Çekirdek Kromozom 5Document62 pagesÇekirdek Kromozom 5Enes BozdağNo ratings yet
- Beslenme Biyokimyası - 1 (Santral Dogma)Document11 pagesBeslenme Biyokimyası - 1 (Santral Dogma)MELİKE CEYDA GÖKTAŞNo ratings yet
- Bakteri̇ Geneti̇ği̇Document58 pagesBakteri̇ Geneti̇ği̇seref100% (1)
- 10-Translasyon Ve Post Translayonel ModifikasyonlarDocument38 pages10-Translasyon Ve Post Translayonel Modifikasyonlargdhvf fbjfghNo ratings yet
- PSİ Biyoloji - ReplikasyonDocument79 pagesPSİ Biyoloji - ReplikasyonYılmaz ŞahinNo ratings yet
- 6 DNA ReplikasyonuDocument65 pages6 DNA Replikasyonugdhvf fbjfghNo ratings yet
- 5-TOKSİKOGENOMİK Ve GENETİK NEDENLERE BAĞLI TOKSİSİTEDocument120 pages5-TOKSİKOGENOMİK Ve GENETİK NEDENLERE BAĞLI TOKSİSİTEŞerif AliNo ratings yet
- Replikasyon 17Document78 pagesReplikasyon 17Halil İbrahim UzunNo ratings yet
- BİY448 Konu-10Document17 pagesBİY448 Konu-10eNo ratings yet
- Pro Gen 10Document79 pagesPro Gen 10Enes BozdağNo ratings yet
- Byl113 Bi̇yoloji̇ 5Document27 pagesByl113 Bi̇yoloji̇ 5subaskavakNo ratings yet
- 12.siinif Biyoloji̇Document48 pages12.siinif Biyoloji̇app77809No ratings yet
- Genomik NGSDocument40 pagesGenomik NGSsehnazcan7No ratings yet
- DNA ReplikasyonuDocument47 pagesDNA ReplikasyonuWissam AlhirakiNo ratings yet
- Endoplazmik Retikulum NotDocument6 pagesEndoplazmik Retikulum NotDuru CanpolatNo ratings yet
- 6 - Hücre Ölümsüzlüğü Ve TümörleşmeDocument74 pages6 - Hücre Ölümsüzlüğü Ve TümörleşmeFurkan YalandanNo ratings yet
- Mitokondri Basic NotesDocument4 pagesMitokondri Basic NotesDuru CanpolatNo ratings yet
- 8) Protei̇n Sentezi̇Document31 pages8) Protei̇n Sentezi̇Hüseyin ArcaNo ratings yet
- 11 DNA'nın ReplikasyonuDocument40 pages11 DNA'nın Replikasyonuesin sakallıNo ratings yet
- NukleusDocument217 pagesNukleusjhgjhNo ratings yet
- Repli Kasy OnDocument77 pagesRepli Kasy Onأحمد دناورNo ratings yet
- RNA Tipleri Ve EkspresyonlarıDocument29 pagesRNA Tipleri Ve EkspresyonlarınurefsansametNo ratings yet
- Epigetiğe Giriş Ve MiRNA'LarDocument62 pagesEpigetiğe Giriş Ve MiRNA'LarTalha KavcarNo ratings yet
- Posttranslasyonel Modifikasyonlar-2019Document99 pagesPosttranslasyonel Modifikasyonlar-2019Serdar ÖzgürNo ratings yet
- Genden Proteine-3 CVPDocument2 pagesGenden Proteine-3 CVPsevdembingollNo ratings yet