You might also like
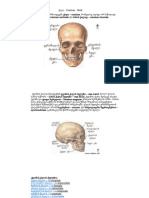
- pbl თავის ქალაDocument30 pagespbl თავის ქალაEvagrius PonticusNo ratings yet
- მიკრობიოლოგიაDocument19 pagesმიკრობიოლოგიაAna ChochishviliNo ratings yet
- 02022022114601 - I- ნაწილი - დოზირებაDocument25 pages02022022114601 - I- ნაწილი - დოზირებაAnush KiNo ratings yet
- მიკრობიოლოგიატესტები2 3Document366 pagesმიკრობიოლოგიატესტები2 3Emil HeydarovNo ratings yet
- სისხლის საერთო ანალიზიDocument2 pagesსისხლის საერთო ანალიზიxvicha0% (1)
- მე 7 შედეგი ბიოქიმია საექთნო განათლებაDocument6 pagesმე 7 შედეგი ბიოქიმია საექთნო განათლებაNati TsiklauriNo ratings yet
- ზოოლოგიის შუალედური 2020Document16 pagesზოოლოგიის შუალედური 2020Tay TaNo ratings yet
- II-შუალედური ზოოლოგიაDocument12 pagesII-შუალედური ზოოლოგიაTay TaNo ratings yet
- პრაქტიკული #9 შემაერთებელი ქსოვილი -savarjishoebitDocument26 pagesპრაქტიკული #9 შემაერთებელი ქსოვილი -savarjishoebitMaria MaisuradzeNo ratings yet
- ფინალური გამოცდა, დავით მაისურაძეDocument5 pagesფინალური გამოცდა, დავით მაისურაძეDavit MaisuradzeNo ratings yet
- გადმოსაწერი ხრიკიDocument8 pagesგადმოსაწერი ხრიკიMari AmiNo ratings yet
- эмбриоDocument9 pagesэмбриоKristinaNo ratings yet
- X Cell, PL MembraneDocument46 pagesX Cell, PL MembraneJohn DoeNo ratings yet
- ბიოლოგიაDocument8 pagesბიოლოგიათათა ბერაძეNo ratings yet
- 1. უჯრედი და გენომიDocument4 pages1. უჯრედი და გენომიMaria MaisuradzeNo ratings yet
- ლექცია 1Document25 pagesლექცია 1Sandro AmiranashviliNo ratings yet
- პათოფიზიოლოგია 1 სემინარიDocument6 pagesპათოფიზიოლოგია 1 სემინარი私の名前は ナナNo ratings yet
- ფინალური ტესტები საყრდენ მამოძრავებელი სისტემაDocument13 pagesფინალური ტესტები საყრდენ მამოძრავებელი სისტემაiza naskidashviliNo ratings yet
- ბიოლოგიაDocument7 pagesბიოლოგიაVano ChokheliNo ratings yet
- ჰისტოლოგია თავი1 2Document84 pagesჰისტოლოგია თავი1 2jasmingiorgadze1986No ratings yet
- Presentation ჰისტოლოგია-სისხლიDocument9 pagesPresentation ჰისტოლოგია-სისხლიShoto TodadzeNo ratings yet
- ნახშირწყლები 1Document62 pagesნახშირწყლები 1Nati TsiklauriNo ratings yet
- ბიოლოგიის საკითხებიDocument7 pagesბიოლოგიის საკითხებიAssyrian PoemsNo ratings yet
- ღვიძლი და ნაღვლის ბუშტიDocument17 pagesღვიძლი და ნაღვლის ბუშტიTika TevdoradzeNo ratings yet
- ბიოქიმია 1 ლქ.1 ნერვ.ქს. ბიოქიმიაDocument31 pagesბიოქიმია 1 ლქ.1 ნერვ.ქს. ბიოქიმიაანა მიქავაNo ratings yet
- ტრანსპორტიDocument28 pagesტრანსპორტიsalome beridze0% (1)
- თერმოდინამიკა ბგერებიDocument47 pagesთერმოდინამიკა ბგერები私の名前は ナナNo ratings yet
- კვებითი ჯაჭვიDocument4 pagesკვებითი ჯაჭვიელენე ჩხიკვაძეNo ratings yet
- 1.ქსოვილები და ჩანასახოვანი ფურცლებიDocument6 pages1.ქსოვილები და ჩანასახოვანი ფურცლებიმაკა გიორგაძეNo ratings yet
- mikrobiologia/მიკრობიოლოგიაDocument3 pagesmikrobiologia/მიკრობიოლოგიაMari AmiNo ratings yet
- ალცჰაიმერიDocument2 pagesალცჰაიმერიgiorgiNo ratings yet
- ლექცია სისხლის ფორმიანი ელემენტებიDocument12 pagesლექცია სისხლის ფორმიანი ელემენტებიNia ChechelashviliNo ratings yet
- გასტრულაცია და ნოტოგენეზიDocument2 pagesგასტრულაცია და ნოტოგენეზიgiorgi0% (1)
- უჯრედის აგებულება და შედგენილობა30Document23 pagesუჯრედის აგებულება და შედგენილობა30Geo-FootballNo ratings yet
- MikroDocument7 pagesMikroNeolina SamushiaNo ratings yet
- მიკრობიოლოგია3 PDFDocument24 pagesმიკრობიოლოგია3 PDFMeduka GelashviliNo ratings yet
- II კვირა -ანატომია (ჯანდაცვა)Document16 pagesII კვირა -ანატომია (ჯანდაცვა)levani metskhvarishviliNo ratings yet
- კონსპექტი (ფინალური)Document81 pagesკონსპექტი (ფინალური)ანი სილაგაძეNo ratings yet
- ბიოლოგია პასუხებიანი 2Document9 pagesბიოლოგია პასუხებიანი 2MariamRevishviliNo ratings yet
- პრაქტიკული 8 - კბილიDocument17 pagesპრაქტიკული 8 - კბილიNin UcaNo ratings yet
- აქტიური ტრანსპორტი CPx ტუმბოების ჩათვლითDocument6 pagesაქტიური ტრანსპორტი CPx ტუმბოების ჩათვლითLika JorbenadzeNo ratings yet
- კონსპექტი 3Document12 pagesკონსპექტი 3Gvantsa MorchadzeNo ratings yet
- ფიზიკა i შუალედურიDocument22 pagesფიზიკა i შუალედურიLasha MomtselidzeNo ratings yet
- პარაზიტოლოგია ენისიDocument43 pagesპარაზიტოლოგია ენისიMariam AbesadzeNo ratings yet
- UntitledDocument16 pagesUntitledMari AzaladzeNo ratings yet
- I-მოდული-გამოტანილები ანატომიაDocument62 pagesI-მოდული-გამოტანილები ანატომიაLika EloshviliNo ratings yet
- ჰისტოს 2 ლექციაDocument10 pagesჰისტოს 2 ლექციაAna UsharidzeNo ratings yet
- ჰეტეროტროფული კვების ტიპები.Document4 pagesჰეტეროტროფული კვების ტიპები.Lika ButikashviliNo ratings yet
- 7. უჯრედული კომუნიკაციებიDocument4 pages7. უჯრედული კომუნიკაციებიGiorgi RakviashviliNo ratings yet
- მიტოქონდრიაDocument4 pagesმიტოქონდრიაnino tsevelidzeNo ratings yet
- HidroDocument34 pagesHidroSaba GabadzeNo ratings yet
- პრაქტიკული 1 ნერვულიDocument37 pagesპრაქტიკული 1 ნერვულიMaria MaisuradzeNo ratings yet
- ბიოლოგია ანატომიაDocument45 pagesბიოლოგია ანატომიაეკა ზამთარაშვილიNo ratings yet
- ბიო შუალედური 1Document18 pagesბიო შუალედური 1Lika EloshviliNo ratings yet
- სამ ბიო 3Document25 pagesსამ ბიო 3kundavid218No ratings yet
- შუალედური გამოცდის ტესტი მოლეკულურ ბიოლოგიაშიDocument12 pagesშუალედური გამოცდის ტესტი მოლეკულურ ბიოლოგიაშიMaghradzee LazareNo ratings yet
- შედეგი 6.მასალა,ტესტები,სტუდენტ.Document24 pagesშედეგი 6.მასალა,ტესტები,სტუდენტ.Natia ShayulashviliNo ratings yet
- ჰისტოლოგიაDocument8 pagesჰისტოლოგიაკოკაNo ratings yet
- ჰისტო ძვლის წარმოქმნაDocument3 pagesჰისტო ძვლის წარმოქმნაKeti KimadzeNo ratings yet
- დიუშენი 2Document11 pagesდიუშენი 2zh6ydczptbNo ratings yet
- ქორდიანებიDocument47 pagesქორდიანებიxvichaNo ratings yet
- სწორი პასუხები - i ვარიანტიDocument1 pageსწორი პასუხები - i ვარიანტიxvichaNo ratings yet
- ლოგიკური მსჯელობა-1Document18 pagesლოგიკური მსჯელობა-1xvichaNo ratings yet
- სწორი პასუხებიDocument1 pageსწორი პასუხებიxvichaNo ratings yet
- GRE Test 2018 LOG CDocument18 pagesGRE Test 2018 LOG CxvichaNo ratings yet
- Cრეაქტიული ცილის განსაზღვრა სისხლის შრატშიDocument1 pageCრეაქტიული ცილის განსაზღვრა სისხლის შრატშიxvichaNo ratings yet
- LR I 2015 GreDocument16 pagesLR I 2015 GrexvichaNo ratings yet
- GRE Test 2018 LOG BDocument19 pagesGRE Test 2018 LOG BxvichaNo ratings yet
- ერითროციტების დალექვის სიჩქარის განსაზღვრა სისხლშიDocument1 pageერითროციტების დალექვის სიჩქარის განსაზღვრა სისხლშიxvichaNo ratings yet
- 2014 Gre Logika lr2 - eDocument16 pages2014 Gre Logika lr2 - exvichaNo ratings yet
- ლოგიკური მსჯელობაDocument18 pagesლოგიკური მსჯელობაxvichaNo ratings yet
- 2014 Gre Gamocdis Testis PasuxebiDocument1 page2014 Gre Gamocdis Testis PasuxebixvichaNo ratings yet
- სწორი პასუხები - c ტიპის ტესტიDocument1 pageსწორი პასუხები - c ტიპის ტესტიxvichaNo ratings yet
- ასდასდDocument19 pagesასდასდxvichaNo ratings yet
- 142155Document3 pages142155xvichaNo ratings yet
- TeodicyDocument31 pagesTeodicyxvichaNo ratings yet
- ქორდიანებიDocument47 pagesქორდიანებიxvichaNo ratings yet
- зplatyhelminthes rotiferaDocument52 pagesзplatyhelminthes rotiferaxvichaNo ratings yet
- $R6OGKYNDocument7 pages$R6OGKYNxvichaNo ratings yet
- ხსნარები, დისოციაცია, კინეტიკაDocument20 pagesხსნარები, დისოციაცია, კინეტიკაxvichaNo ratings yet