You might also like
- სისხლის საერთო ანალიზიDocument2 pagesსისხლის საერთო ანალიზიxvicha0% (1)
- თინათინ გაბაძეDocument2 pagesთინათინ გაბაძეTika GabadzeNo ratings yet
- მცენარეთა განოყიერების სისტემებიDocument73 pagesმცენარეთა განოყიერების სისტემებიNikaJafiashviliNo ratings yet
- Á Á Á Á Á Á Á Á 8 Á Á Á Á Á Á Á Á Á Á Á ÁDocument17 pagesÁ Á Á Á Á Á Á Á 8 Á Á Á Á Á Á Á Á Á Á Á Ágiorgi.gvaladze.05No ratings yet
- ლექცია 5 ფესვი, ყლორტი, ღეროDocument5 pagesლექცია 5 ფესვი, ყლორტი, ღეროSofia KutsenkoNo ratings yet
- ყლორტი,ღერო. მორფოლოგია, ანატომია, მეტამორფოზებიDocument9 pagesყლორტი,ღერო. მორფოლოგია, ანატომია, მეტამორფოზებინიკოლოზ გიორბელიძეNo ratings yet
- მევენახეობა 4Document15 pagesმევენახეობა 4Mariam GurgenishviliNo ratings yet
- თემა 8 3Document7 pagesთემა 8 3MariiNo ratings yet
- Á Á Á Á Á Á Á Á 2 Á ¡Á Á ¡Á Á Á Á Á ®á Á Á Á Á Á Á Á Á 3Document7 pagesÁ Á Á Á Á Á Á Á 2 Á ¡Á Á ¡Á Á Á Á Á ®á Á Á Á Á Á Á Á Á 3giorgi.gvaladze.05No ratings yet
- ფოთოლიDocument16 pagesფოთოლიLevan ZaalishviliNo ratings yet
- FesviDocument4 pagesFesviDima SajaiaNo ratings yet
- ფოთოლიDocument14 pagesფოთოლიmariami.khmiadashviliNo ratings yet
- ვაშლის მოვლა-მოყვანის ტექნოლოგიაDocument6 pagesვაშლის მოვლა-მოყვანის ტექნოლოგიაGaga ChubinidzeNo ratings yet
- ტოლსპორიანი გვიმრების გამრავლებაDocument4 pagesტოლსპორიანი გვიმრების გამრავლებაLevan ZaalishviliNo ratings yet
- ფოთოლიDocument14 pagesფოთოლიmariami.khmiadashviliNo ratings yet
- ლექცია 10 სამეფო Protista - წყალმცენარეები, გამრავლება -გავრცელება, ეკოლოგია,Document17 pagesლექცია 10 სამეფო Protista - წყალმცენარეები, გამრავლება -გავრცელება, ეკოლოგია,Sofia KutsenkoNo ratings yet
- N 9 ხავსებიDocument4 pagesN 9 ხავსებიluka inasaridzeNo ratings yet
- ჟოლო - სასწავლო მასალა შემოკლებულიDocument9 pagesჟოლო - სასწავლო მასალა შემოკლებულიZviad BobokashviliNo ratings yet
- შიშველთესლოვნებიDocument2 pagesშიშველთესლოვნებიnika j0% (2)
- ყვავილოვან მცენარეთა განაყოფიერებაDocument7 pagesყვავილოვან მცენარეთა განაყოფიერებაluka chumburidze100% (2)
- თხილის მოვლა-მოყვანის ტექნოლოგიაDocument4 pagesთხილის მოვლა-მოყვანის ტექნოლოგიაGaga ChubinidzeNo ratings yet
- UntitledDocument7 pagesUntitledანა ხათრიძეNo ratings yet
- სასათბურე მეურნეობა პროექტი მარიამ თენოშვილი. IV კურსი.Document21 pagesსასათბურე მეურნეობა პროექტი მარიამ თენოშვილი. IV კურსი.aiase2022No ratings yet
- ხავსიDocument1 pageხავსიmdnmfhyc48No ratings yet
- მცნ.ფიზიოლოგიის რეფერატიDocument5 pagesმცნ.ფიზიოლოგიის რეფერატიtamta bochorishviliNo ratings yet
- Biologia Evolucia Seguebulobna Da Aseshemdeg'Document2 pagesBiologia Evolucia Seguebulobna Da Aseshemdeg'kvitaantonianiNo ratings yet
- Á Á Á Á Á Á Á Á 5 Á Á Á Á Á Á Á Á Á Á Á Á Á ÁDocument9 pagesÁ Á Á Á Á Á Á Á 5 Á Á Á Á Á Á Á Á Á Á Á Á Á Ágiorgi.gvaladze.05No ratings yet
- 2. ბიოდეგრადაციაDocument10 pages2. ბიოდეგრადაციაMariam ShoniaNo ratings yet
- Chapter 3 2022 Recombinantion of GenesDocument25 pagesChapter 3 2022 Recombinantion of Genesდათა მერებაშვილიNo ratings yet
- ვაზი PDFDocument6 pagesვაზი PDFLasha KvaraNo ratings yet
- ვაზი PDFDocument6 pagesვაზი PDFdeminademina1995No ratings yet
- ვაზი PDFDocument6 pagesვაზი PDFIrma IrmaNo ratings yet
- სამკურნალოDocument4 pagesსამკურნალოtamta bochorishviliNo ratings yet
- 7 DamulchvaDocument6 pages7 DamulchvaMKGUNo ratings yet
- USAID NEO Apple Guidebook 88541 PDFDocument86 pagesUSAID NEO Apple Guidebook 88541 PDFNikaJafiashviliNo ratings yet
- ბიოDocument3 pagesბიოBadriNo ratings yet
- საკვები კულტურები - შესავალი მდელოდ საკვებწარმოებაშიDocument145 pagesსაკვები კულტურები - შესავალი მდელოდ საკვებწარმოებაშიNikaJafiashviliNo ratings yet
- ენტო ფინალურიDocument7 pagesენტო ფინალურიLuka TkeshelashviliNo ratings yet
- დეკორც მებაღ.მე-5 ლექც.Document24 pagesდეკორც მებაღ.მე-5 ლექც.ლაშა ხმალაძე100% (1)
- ბოტანიკა შუალედური studentebs2Document46 pagesბოტანიკა შუალედური studentebs2Giorgi AvalianiNo ratings yet
- ბოტანიკა შუალედური studentebs2Document46 pagesბოტანიკა შუალედური studentebs2Giorgi AvalianiNo ratings yet
- ტუბერ. და ძირნაყ. კულტ.Document5 pagesტუბერ. და ძირნაყ. კულტ.aniNo ratings yet
- 18 MsxaliDocument6 pages18 MsxalikhoteuriNo ratings yet
- BotanikaDocument7 pagesBotanikaნუზაNo ratings yet
- ვაზის სიყვითლეები.Document27 pagesვაზის სიყვითლეები.Mariam GurgenishviliNo ratings yet
- მავნებლები 1Document29 pagesმავნებლები 1Mate LiluashviliNo ratings yet
- ლექცია-პრაქტიკული 7Document3 pagesლექცია-პრაქტიკული 7ნიკა დუშუაშვილიNo ratings yet
- 393Document5 pages393Mariam TakashviliNo ratings yet
- X Cell, PL MembraneDocument46 pagesX Cell, PL MembraneJohn DoeNo ratings yet
- ანატომიაDocument48 pagesანატომიაKeti ShalikashviliNo ratings yet
- AgkiDocument12 pagesAgkiketusha bakradzeNo ratings yet
- უჯრედიDocument13 pagesუჯრედიAnastasia GoduadzeNo ratings yet
- პომიდვრის აგროტექნოლოგიაDocument8 pagesპომიდვრის აგროტექნოლოგიაMisha JaparidzeNo ratings yet
- ფარულთესლოვნებიDocument4 pagesფარულთესლოვნებიnika j100% (1)
- Platyhelminthes 2020Document19 pagesPlatyhelminthes 2020xvichaNo ratings yet
- ლექცია 3.სოკოები აგებულება, კლასიფიკაციაDocument23 pagesლექცია 3.სოკოები აგებულება, კლასიფიკაციაtamarkharshiladze2No ratings yet
- 4Document15 pages4ketusha bakradzeNo ratings yet
- უჯრედის ონტოგენეზიDocument20 pagesუჯრედის ონტოგენეზიAnaNo ratings yet
- კვებითი ჯაჭვიDocument4 pagesკვებითი ჯაჭვიელენე ჩხიკვაძეNo ratings yet
- სწორი პასუხები - i ვარიანტიDocument1 pageსწორი პასუხები - i ვარიანტიxvichaNo ratings yet
- ერითროციტების დალექვის სიჩქარის განსაზღვრა სისხლშიDocument1 pageერითროციტების დალექვის სიჩქარის განსაზღვრა სისხლშიxvichaNo ratings yet
- სწორი პასუხებიDocument1 pageსწორი პასუხებიxvichaNo ratings yet
- ლოგიკური მსჯელობა-1Document18 pagesლოგიკური მსჯელობა-1xvichaNo ratings yet
- Cრეაქტიული ცილის განსაზღვრა სისხლის შრატშიDocument1 pageCრეაქტიული ცილის განსაზღვრა სისხლის შრატშიxvichaNo ratings yet
- GRE Test 2018 LOG CDocument18 pagesGRE Test 2018 LOG CxvichaNo ratings yet
- 2014 Gre Logika lr2 - eDocument16 pages2014 Gre Logika lr2 - exvichaNo ratings yet
- ლოგიკური მსჯელობაDocument18 pagesლოგიკური მსჯელობაxvichaNo ratings yet
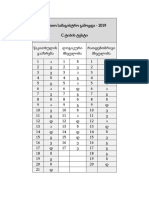
- სწორი პასუხები - c ტიპის ტესტიDocument1 pageსწორი პასუხები - c ტიპის ტესტიxvichaNo ratings yet
- 142155Document3 pages142155xvichaNo ratings yet
- LR I 2015 GreDocument16 pagesLR I 2015 GrexvichaNo ratings yet
- GRE Test 2018 LOG BDocument19 pagesGRE Test 2018 LOG BxvichaNo ratings yet
- 2014 Gre Gamocdis Testis PasuxebiDocument1 page2014 Gre Gamocdis Testis PasuxebixvichaNo ratings yet
- ასდასდDocument19 pagesასდასდxvichaNo ratings yet
- TeodicyDocument31 pagesTeodicyxvichaNo ratings yet
- Platyhelminthes 2020Document19 pagesPlatyhelminthes 2020xvichaNo ratings yet
- ქორდიანებიDocument47 pagesქორდიანებიxvichaNo ratings yet
- зplatyhelminthes rotiferaDocument52 pagesзplatyhelminthes rotiferaxvichaNo ratings yet
- ხსნარები, დისოციაცია, კინეტიკაDocument20 pagesხსნარები, დისოციაცია, კინეტიკაxvichaNo ratings yet