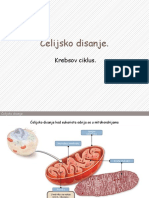
You might also like
- Krebsov Ciklus. Elektron - Transportni LanacDocument18 pagesKrebsov Ciklus. Elektron - Transportni LanacGoranNo ratings yet
- Biologija - Početne LekcijeDocument10 pagesBiologija - Početne LekcijeLazar Ljubenović100% (3)
- Tehnologija Vode Vezbe11 Jonska Izmena 14052014Document4 pagesTehnologija Vode Vezbe11 Jonska Izmena 14052014ИванаСтанићNo ratings yet
- CitologijaDocument24 pagesCitologijaLjubica Nikolic100% (1)
- Predavanje 1Document40 pagesPredavanje 1FotokopirnicaCopyStudioNiksicNo ratings yet
- Fiziologija IspitDocument106 pagesFiziologija IspitNemanja Markovic0% (1)
- Fotosinteza, I Transpiracija 6.razredDocument11 pagesFotosinteza, I Transpiracija 6.razredMilica PetkovićNo ratings yet
- Интрацелуларна и екстрацелуларна срединаDocument14 pagesИнтрацелуларна и екстрацелуларна срединаIlija 5000No ratings yet
- Fotosinteza I Hemosinteza PrezentacijaDocument20 pagesFotosinteza I Hemosinteza PrezentacijaСлађана Арсенић67% (3)
- ЕнзимиDocument8 pagesЕнзимиTijana ZorklijaNo ratings yet
- Fizika Membrane I Nervnog ImpulsaDocument38 pagesFizika Membrane I Nervnog Impulsasladjad83No ratings yet
- Mineralna IshranaDocument93 pagesMineralna IshranaMirjana BugarinNo ratings yet
- 19 BubregDocument7 pages19 BubregTamara StjepovicNo ratings yet
- УСВАЈАЊЕ ВОДЕ И МИНЕРАЛА, ТРАНСПИРАЦИЈА И ТРАНСПОРТDocument20 pagesУСВАЈАЊЕ ВОДЕ И МИНЕРАЛА, ТРАНСПИРАЦИЈА И ТРАНСПОРТSuzana SotirovNo ratings yet
- I Nedelja PredavanjaDocument25 pagesI Nedelja PredavanjasrdjanNo ratings yet
- Фотосинтеза-Уна СупчинDocument26 pagesФотосинтеза-Уна СупчинUna SurcinNo ratings yet
- Beleske Iz Hemije 4. RazredDocument16 pagesBeleske Iz Hemije 4. RazrednekokocitalektiruNo ratings yet
- Hemija 3009Document9 pagesHemija 3009nekokocitalektiruNo ratings yet
- Dinamika Bioloških MembranaDocument10 pagesDinamika Bioloških MembranaMiki MausNo ratings yet
- II KOLOKVIJUM IZ BIOFIZIKE - Fizika Membrane I Nervnog Impulsa Biomehanicki Procesi 1) Fizika Membrane I Nervnog ImpulsaDocument11 pagesII KOLOKVIJUM IZ BIOFIZIKE - Fizika Membrane I Nervnog Impulsa Biomehanicki Procesi 1) Fizika Membrane I Nervnog ImpulsaAleksandra MilicevicNo ratings yet
- 8. FOTOSINTEZA (1)Document59 pages8. FOTOSINTEZA (1)kjunackovNo ratings yet
- Од Прве Ћелије До ЕукариотаDocument16 pagesОд Прве Ћелије До ЕукариотаSofija PetrovicNo ratings yet
- Lanac Disanja5Document26 pagesLanac Disanja5Rastko ZečevićNo ratings yet
- BiologijaDocument5 pagesBiologijaTamara MitrovićNo ratings yet
- Funkcija Bioloskih Membrana - Endozom - Lizozom 02Document26 pagesFunkcija Bioloskih Membrana - Endozom - Lizozom 02Mirjana PrekovicNo ratings yet
- Biologija - DomaćiDocument2 pagesBiologija - DomaćiMarijaaaaNo ratings yet
- BiosDocument2 pagesBiosLazar MedarNo ratings yet
- 02 CelijaDocument76 pages02 CelijaanimeNo ratings yet
- 7 Transport Nutrijenata CelijskiADocument63 pages7 Transport Nutrijenata CelijskiAMirjana MihajlovićNo ratings yet
- Svijetla I Tamna Faza FotosintezeDocument38 pagesSvijetla I Tamna Faza FotosintezeMinela RidžalNo ratings yet
- Gajton Celijeske MembraneDocument12 pagesGajton Celijeske MembraneSlavenNo ratings yet
- Osmoza I DifuzijaDocument16 pagesOsmoza I DifuzijaMiks MaksNo ratings yet
- Hemija 0125Document35 pagesHemija 0125nekokocitalektiruNo ratings yet
- Aktivan Transport Vode I Mineralnih MaterijaDocument15 pagesAktivan Transport Vode I Mineralnih MaterijaMilan GoranovićNo ratings yet
- Fizio IIDocument16 pagesFizio IIMiljana PetrovicNo ratings yet
- Fotos in TezaDocument7 pagesFotos in TezaНенад ПавловићNo ratings yet
- Membrana Transport EndocitozaDocument26 pagesMembrana Transport EndocitozaBag of CornNo ratings yet
- BIOGENI ELEMENTI Ucetsvuju U Gradji Zivih Organizama: - H2CO3 H+ + HCO3-slabo DisosujeDocument35 pagesBIOGENI ELEMENTI Ucetsvuju U Gradji Zivih Organizama: - H2CO3 H+ + HCO3-slabo DisosujenekokocitalektiruNo ratings yet
- CITOLOGIJADocument71 pagesCITOLOGIJAelmedina omeragicNo ratings yet
- Ćelijske Membrane I Transport-Prevod OP Iz BiohemijeDocument8 pagesĆelijske Membrane I Transport-Prevod OP Iz BiohemijeAmar MaseticNo ratings yet
- Transport Kroz MembranuDocument24 pagesTransport Kroz MembranuMajaNo ratings yet
- ПИТАЊА ИЗ БИОЛОГИЈЕDocument3 pagesПИТАЊА ИЗ БИОЛОГИЈЕteoNo ratings yet
- 10 MetabolizamDocument24 pages10 MetabolizamTodorov.TodorNo ratings yet
- Fiziologija Celijske MembraneDocument54 pagesFiziologija Celijske MembraneNenad ĐorđevićNo ratings yet
- Kardiovaskularni Sistem - Smer EkologijaDocument37 pagesKardiovaskularni Sistem - Smer EkologijaAlex AnnieNo ratings yet
- Mikrobni Enzimi Word 3Document66 pagesMikrobni Enzimi Word 3Miroslav Cici PavelkicNo ratings yet
- Hemijski Sastav ĆelijeDocument34 pagesHemijski Sastav ĆelijeedinNo ratings yet
- Metan U VodiDocument8 pagesMetan U VoditihatikNo ratings yet
- Celija I BiomolekuliDocument72 pagesCelija I BiomolekuliFilip PlavinacNo ratings yet
- Untitled Document-4 PDFDocument4 pagesUntitled Document-4 PDFVanja TomicNo ratings yet
- Fiziologija Kolokvijum IIDocument5 pagesFiziologija Kolokvijum IIМарко СтевановићNo ratings yet
- Vodni Rezim 2018 Milan I BobaDocument67 pagesVodni Rezim 2018 Milan I BobaEd NikolicNo ratings yet
- Promet Kroz Celijsku MembranuDocument33 pagesPromet Kroz Celijsku MembranuIskra DejanovicNo ratings yet
- ЕНЗИМИDocument22 pagesЕНЗИМИŽeljko MirkovićNo ratings yet
- Bioloske MolekuleDocument22 pagesBioloske MolekuleMartina GluhanNo ratings yet
- ПИТАЊА ЗА ГОДИШЊИ ТЕСТ ИЗ БИОЛОГИЈЕ ЗА ПРВИ РАЗРЕДDocument4 pagesПИТАЊА ЗА ГОДИШЊИ ТЕСТ ИЗ БИОЛОГИЈЕ ЗА ПРВИ РАЗРЕДJasminaRedzovicNo ratings yet