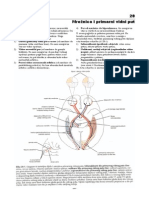
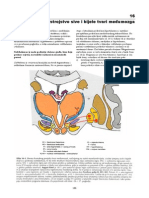
You might also like
- NeurologijaDocument81 pagesNeurologijaIva Ladić100% (4)
- NEUROLOGIJADocument137 pagesNEUROLOGIJAktrokanovicNo ratings yet
- Skripta Iz Temelja NeuroznanostiDocument118 pagesSkripta Iz Temelja NeuroznanostiperolegendaNo ratings yet
- Biologija-Osjetilni I Zivcani SustavDocument5 pagesBiologija-Osjetilni I Zivcani SustavFilipIvanovicNo ratings yet
- P35 - Mali MozakDocument8 pagesP35 - Mali MozakErikaNo ratings yet
- Živčani SustavDocument16 pagesŽivčani SustavmirelaNo ratings yet
- Skripta - 1Document15 pagesSkripta - 1Duje RazmilovićNo ratings yet
- Temelji Neuroznanosti - 35 PoglavljeDocument9 pagesTemelji Neuroznanosti - 35 PoglavljeAndika HattersplatNo ratings yet
- Skripta Završni IspitDocument57 pagesSkripta Završni IspitAlija AleNo ratings yet
- CNS 1Document4 pagesCNS 1Selma MuratovicNo ratings yet
- Cns FarmacijaDocument11 pagesCns FarmacijaAmira RanicaNo ratings yet
- Neuronske MrežeDocument21 pagesNeuronske MrežeAmira TrakoNo ratings yet
- Crchap 7Document3 pagesCrchap 7Edita ZaimovicNo ratings yet
- Anatomija 2polDocument42 pagesAnatomija 2polDorotea BratekNo ratings yet
- Teodora - Karalic - Seminarski - Prepravljen - Docx Filename UTF-8''teodora Karalic Seminarski PrepravljenDocument3 pagesTeodora - Karalic - Seminarski - Prepravljen - Docx Filename UTF-8''teodora Karalic Seminarski PrepravljenSinisa Ristic100% (1)
- Mali Mozak Uvodni Dio Marko UljatDocument17 pagesMali Mozak Uvodni Dio Marko UljatGoran SimicNo ratings yet
- Neurologija 1Document0 pagesNeurologija 1SomeoneNo ratings yet
- Kemijska Neurotransmisija - Stahl's Essential Psychopharmacology - Neuroscientific Basis and Practical Applications, 4th EdDocument41 pagesKemijska Neurotransmisija - Stahl's Essential Psychopharmacology - Neuroscientific Basis and Practical Applications, 4th EdDragutin PetrićNo ratings yet
- NB Neuropsihologija Druga ParcijalaDocument20 pagesNB Neuropsihologija Druga ParcijalaMiran KosticNo ratings yet
- IUS 2.dio SKRIPTA PDFDocument163 pagesIUS 2.dio SKRIPTA PDFAdnan BabićNo ratings yet
- Fiziologija CnsDocument102 pagesFiziologija CnsSulejman HadzicNo ratings yet
- Struktura I Funkcija Neurona HANDOUT Osnove Biološke PsihologijeDocument5 pagesStruktura I Funkcija Neurona HANDOUT Osnove Biološke PsihologijeLana OstojićNo ratings yet
- BIOLOGIJADocument10 pagesBIOLOGIJAdarko horvatNo ratings yet
- Biologija Drugo IspitivanjeDocument12 pagesBiologija Drugo IspitivanjeivonaNo ratings yet
- CNSDocument43 pagesCNSprsno100% (1)
- Živčani SustavDocument7 pagesŽivčani SustavDorotea ProsnikNo ratings yet
- Skripta 1 MergedDocument226 pagesSkripta 1 MergedInstrukcije SeminariNo ratings yet
- Neuronske MrezeDocument26 pagesNeuronske MrezeJanko StrusaNo ratings yet
- Skripta Iz TNZa PDFDocument130 pagesSkripta Iz TNZa PDFApoteka LjiljanNo ratings yet
- Temelji Neuroznanosti - 28 PoglavljeDocument18 pagesTemelji Neuroznanosti - 28 PoglavljeAndika HattersplatNo ratings yet
- Poglavlje 56, P10Document6 pagesPoglavlje 56, P10Ena PaparićNo ratings yet
- Živčani Sustav - 1.dioDocument3 pagesŽivčani Sustav - 1.dioMartina KneževićNo ratings yet
- Neurologija - Skripta (HR)Document31 pagesNeurologija - Skripta (HR)olegNo ratings yet
- Temelji Neuroznanosti - 10 PoglavljeDocument17 pagesTemelji Neuroznanosti - 10 PoglavljeAndika HattersplatNo ratings yet
- Biologija PitanjaDocument4 pagesBiologija PitanjaivonaNo ratings yet
- Javor Vouk-Kamenski - Tema 1. AntidepresiviDocument29 pagesJavor Vouk-Kamenski - Tema 1. AntidepresiviJavor Vouk-KamenskiNo ratings yet
- Temelji Neuroznanosti - 37 PoglavljeDocument10 pagesTemelji Neuroznanosti - 37 PoglavljeAndika HattersplatNo ratings yet
- Temelji Neuroznanosti - 33 PoglavljeDocument10 pagesTemelji Neuroznanosti - 33 PoglavljeAndika HattersplatNo ratings yet
- Temelji Neuroznanosti - 34 PoglavljeDocument8 pagesTemelji Neuroznanosti - 34 PoglavljeAndika HattersplatNo ratings yet
- Temelji Neuroznanosti - 32 PoglavljeDocument12 pagesTemelji Neuroznanosti - 32 PoglavljeAndika HattersplatNo ratings yet
- Temelji Neuroznanosti - 29 PoglavljeDocument14 pagesTemelji Neuroznanosti - 29 PoglavljeAndika HattersplatNo ratings yet
- Temelji Neuroznanosti - 27 PoglavljeDocument17 pagesTemelji Neuroznanosti - 27 PoglavljeAndika HattersplatNo ratings yet
- Temelji Neuroznanosti - 22 PoglavljeDocument6 pagesTemelji Neuroznanosti - 22 PoglavljeAndika HattersplatNo ratings yet
- Temelji Neuroznanosti - 28 PoglavljeDocument18 pagesTemelji Neuroznanosti - 28 PoglavljeAndika HattersplatNo ratings yet
- Temelji Neuroznanosti - 23 PoglavljeDocument7 pagesTemelji Neuroznanosti - 23 PoglavljeAndika HattersplatNo ratings yet
- Temelji Neuroznanosti - 18 PoglavljeDocument12 pagesTemelji Neuroznanosti - 18 PoglavljeAndika HattersplatNo ratings yet
- Temelji Neuroznanosti - 20 PoglavljeDocument11 pagesTemelji Neuroznanosti - 20 PoglavljeAndika Hattersplat100% (1)
- Temelji Neuroznanosti - 16 PoglavljeDocument6 pagesTemelji Neuroznanosti - 16 PoglavljeAndika HattersplatNo ratings yet
- Temelji Neuroznanosti - 02 PoglavljeDocument10 pagesTemelji Neuroznanosti - 02 PoglavljeDobroša RamaNo ratings yet
- Temelji Neuroznanosti - 10 PoglavljeDocument17 pagesTemelji Neuroznanosti - 10 PoglavljeAndika HattersplatNo ratings yet
- Temelji Neuroznanosti - 05 PoglavljeDocument21 pagesTemelji Neuroznanosti - 05 PoglavljeAndika HattersplatNo ratings yet
- Temelji Neuroznanosti - 01 PoglavljeDocument4 pagesTemelji Neuroznanosti - 01 PoglavljeTea OmanovicNo ratings yet