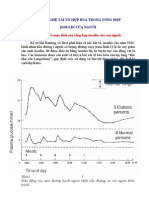
You might also like
- Mô hình kinh doanh thức uống trà Kombucha - Môn quản lý dịch vụ ẩm thựcDocument76 pagesMô hình kinh doanh thức uống trà Kombucha - Môn quản lý dịch vụ ẩm thựcNam NguyenHoangNo ratings yet
- Xúc tác sinh họcDocument11 pagesXúc tác sinh họcyenphuongngocnNo ratings yet
- Case CHNLDocument8 pagesCase CHNLPhương PhanNo ratings yet
- 44-Article Text-57-1-10-20210915 PDFDocument51 pages44-Article Text-57-1-10-20210915 PDFNguyễn TrườngNo ratings yet
- Sinh Lý Thực VậtDocument3 pagesSinh Lý Thực Vật[•Bé Oanh•]No ratings yet
- 02. TL - CÂU HỎI ÔN TẬP HOÁ DƯỢC 01Document6 pages02. TL - CÂU HỎI ÔN TẬP HOÁ DƯỢC 01dkduy-duoc16No ratings yet
- CiprofloxacinDocument9 pagesCiprofloxacinThư HuỳnhNo ratings yet
- ĐC Hóa SinhDocument14 pagesĐC Hóa SinhDũng HoàngNo ratings yet
- Phan Ung Nau Hoa Cua EnzymeDocument7 pagesPhan Ung Nau Hoa Cua EnzymeAnh QuỳnhNo ratings yet
- Hóa SinhDocument46 pagesHóa Sinhhangminhkk2277No ratings yet
- COPDDocument3 pagesCOPDDũng NguyễnNo ratings yet
- Q&A Chương 3 (Tế Bào Học)Document4 pagesQ&A Chương 3 (Tế Bào Học)Kessy KessyNo ratings yet
- Case ReportDocument10 pagesCase ReportAnh ĐoànNo ratings yet
- Chương 79.Insulin, glucagon, và đái tháo đường - Hoàng NguyễnDocument33 pagesChương 79.Insulin, glucagon, và đái tháo đường - Hoàng NguyễnDương DươngNo ratings yet
- 2 hợp chất kháng viêmDocument6 pages2 hợp chất kháng viêmThảo PhươngNo ratings yet
- Tình trạng thiếu oxy âm thầm trong COVID 19Document6 pagesTình trạng thiếu oxy âm thầm trong COVID 19Alex NgNo ratings yet
- Hóa Dư CDocument17 pagesHóa Dư Cngoc anh doanNo ratings yet
- SINH LÍ CÂN BẰNG NƯỚC VÀ CÁC CHẤT ĐIỆN GIẢI VTCDocument6 pagesSINH LÍ CÂN BẰNG NƯỚC VÀ CÁC CHẤT ĐIỆN GIẢI VTCQuốc NguyễnNo ratings yet
- Bài Giảng Hóa Sinh Hệ Tiết NiệuDocument61 pagesBài Giảng Hóa Sinh Hệ Tiết NiệuVân anh NguyễnNo ratings yet
- ĐỀ CƯƠNG ÔN TẬP MÔN HÓA DƯỢC 2Document15 pagesĐỀ CƯƠNG ÔN TẬP MÔN HÓA DƯỢC 2trangb2004234No ratings yet
- QUINOLONEDocument17 pagesQUINOLONENguyễn HàNo ratings yet
- VIE-VIÊM TỤY CẤP-finalDocument4 pagesVIE-VIÊM TỤY CẤP-finalJungjackNo ratings yet
- #Asthma #Spirometry #Inflammation #Cytokines #ChemokinesDocument20 pages#Asthma #Spirometry #Inflammation #Cytokines #Chemokines2151010465No ratings yet
- 1.máY TRUYỀN DỊCHDocument15 pages1.máY TRUYỀN DỊCHKhoa NhiNo ratings yet
- INSULIN VÀ THUỐC HẠ ĐƯỜNG HUYẾTDocument14 pagesINSULIN VÀ THUỐC HẠ ĐƯỜNG HUYẾTdigoxin_y34No ratings yet
- Giới thiệu Enzyme AsparanaseDocument13 pagesGiới thiệu Enzyme AsparanaseNhật Ngô ThanhNo ratings yet
- 21 ĐIỀU TRỊ THIẾU MÁU Ở BỆNH NHÂN SUY THẬN MẠNDocument11 pages21 ĐIỀU TRỊ THIẾU MÁU Ở BỆNH NHÂN SUY THẬN MẠNcuong voNo ratings yet
- Khi NoDocument7 pagesKhi NoNgô Thùy AnNo ratings yet
- Chuyển Hóa ProteinDocument42 pagesChuyển Hóa ProteinMy KongNo ratings yet
- Su Ho Hap Cua Te BaoDocument19 pagesSu Ho Hap Cua Te BaoThi Võ HồngNo ratings yet
- Vận chuyển đường và Bệnh tiểu đường liên quan tới GLUTDocument29 pagesVận chuyển đường và Bệnh tiểu đường liên quan tới GLUTNguyễn Thanh ThảoNo ratings yet
- SLB Đái Tháo Đư NGDocument13 pagesSLB Đái Tháo Đư NGtramht224022snNo ratings yet
- Bệnh học: BÀI 2 ĐẠI CƯƠNG VỀ BL TIẾT NIỆUDocument20 pagesBệnh học: BÀI 2 ĐẠI CƯƠNG VỀ BL TIẾT NIỆUwy linhNo ratings yet
- Đại cương về thuốc ức chế men chuyểnDocument10 pagesĐại cương về thuốc ức chế men chuyểnLiên ĐàoNo ratings yet
- xét nghiệm proteinDocument7 pagesxét nghiệm proteinNguyen Duy Tien B2010797No ratings yet
- Hóa học Acid NucleicDocument67 pagesHóa học Acid NucleicVân NguyễnNo ratings yet
- 2022-Sinh Lý Bệnh Hô HấpDocument17 pages2022-Sinh Lý Bệnh Hô HấpThị Hoàn LêNo ratings yet
- DNA Tái T H P T o InsulinDocument11 pagesDNA Tái T H P T o InsulinNgo Tien DatNo ratings yet
- Sinh Lý MáuDocument30 pagesSinh Lý Máunguyenngocthu.workNo ratings yet
- HOÁ HỌC PROTEIN PDFDocument54 pagesHOÁ HỌC PROTEIN PDFTrang Nguyễn50% (2)
- 2021-SINH LÝ BỆNH HÔ HẤP-sau thông quaDocument17 pages2021-SINH LÝ BỆNH HÔ HẤP-sau thông quaduy bùiNo ratings yet
- Đề 15 nhóm k18Document20 pagesĐề 15 nhóm k18Phan Như RiNo ratings yet
- CH 10 Carbohydrates-Môn hóa sinh họcDocument61 pagesCH 10 Carbohydrates-Môn hóa sinh họcNam NguyenHoang100% (1)
- BÁO CÁO HÓA SINH Nhóm 6Document23 pagesBÁO CÁO HÓA SINH Nhóm 6Ivy TranNo ratings yet
- Bài 25 - Hô Hấp Tế BàoDocument11 pagesBài 25 - Hô Hấp Tế Bàoquoc.70212095tpe2No ratings yet
- LactateDocument18 pagesLactateHanh NguyenNo ratings yet
- Đ I Cương NucleotideDocument26 pagesĐ I Cương NucleotideNgọc CaiNo ratings yet
- Case Lâm Sàng TT Dư C Lý 2Document4 pagesCase Lâm Sàng TT Dư C Lý 2odghvty221120No ratings yet
- Hóa SinhDocument5 pagesHóa SinhNguyễn Hàn LâmNo ratings yet
- câu hỏi ôn tập vi sinh cơ sởDocument10 pagescâu hỏi ôn tập vi sinh cơ sởThành TrịnhNo ratings yet
- RỐI LOẠN CHUYỂN HÓA GLUCIDDocument32 pagesRỐI LOẠN CHUYỂN HÓA GLUCIDVy HảiNo ratings yet
- SINH HỌC DI TRUYỀNDocument8 pagesSINH HỌC DI TRUYỀNHoang Nguyen Anh ThuNo ratings yet
- FILE - 20220614 - 090153 - Chuyen Hoa AcidaminDocument15 pagesFILE - 20220614 - 090153 - Chuyen Hoa Acidaminphuongahiru2303No ratings yet
- TV VLTL HH Nhi NG D NGDocument62 pagesTV VLTL HH Nhi NG D NGThanh NguyenNo ratings yet
- (123doc) - Cac-Qua-Trinh-Bien-Doi-Cua-Thit-Sau-Giet-MoDocument32 pages(123doc) - Cac-Qua-Trinh-Bien-Doi-Cua-Thit-Sau-Giet-Mothanh nguyenNo ratings yet
- Bài 20 PL5 Liệu Pháp Ôxy Ths Trang-1Document19 pagesBài 20 PL5 Liệu Pháp Ôxy Ths Trang-1thuylan11102005No ratings yet
- 18150267-Do Thi My Xuan-Duoc Ly 1-01Document11 pages18150267-Do Thi My Xuan-Duoc Ly 1-01doNo ratings yet
- B - 11.5 - Drug - Metabolism - Revised - 1Document46 pagesB - 11.5 - Drug - Metabolism - Revised - 1Dương LinhNo ratings yet
- Uống nước sinh tố: Phương pháp kỳ diệu - bảo vệ sức khỏe và trị liệu bệnh tật.From EverandUống nước sinh tố: Phương pháp kỳ diệu - bảo vệ sức khỏe và trị liệu bệnh tật.No ratings yet
- Xây dựng hệ thống Haccp- Snack khoai tây vị phô mai KracksDocument15 pagesXây dựng hệ thống Haccp- Snack khoai tây vị phô mai KracksNam NguyenHoangNo ratings yet
- Tổng hợp kiến thức về di truyền học- https://suckhoeditruyen.vnDocument67 pagesTổng hợp kiến thức về di truyền học- https://suckhoeditruyen.vnNam NguyenHoang100% (1)
- 2021 Mau Khoa Luan Tot NghiepDocument19 pages2021 Mau Khoa Luan Tot NghiepGiang Phạm HuỳnhNo ratings yet
- Vai trò của vận động trong quản lý thừa cân béo phì- Học thuật với chuyên gia lần 2Document59 pagesVai trò của vận động trong quản lý thừa cân béo phì- Học thuật với chuyên gia lần 2Nam NguyenHoangNo ratings yet
- Bài báo cáo thực hành môn công nghệ chế biến nông sản- IUHDocument35 pagesBài báo cáo thực hành môn công nghệ chế biến nông sản- IUHNam NguyenHoangNo ratings yet
- Chương 3- Môn thiết kế và phát triển sản phẩm - IUHDocument27 pagesChương 3- Môn thiết kế và phát triển sản phẩm - IUHNam NguyenHoangNo ratings yet
- Tổng Quan Các Chất Độc Acrylamide, PAHs, Furan Trong Chiên Nướng Từ Các Sản Phẩm Tinh Bột- IUHDocument60 pagesTổng Quan Các Chất Độc Acrylamide, PAHs, Furan Trong Chiên Nướng Từ Các Sản Phẩm Tinh Bột- IUHNam NguyenHoangNo ratings yet
- Chương 5 - Môn thiết kế và phát triển sản phẩm - IUHDocument11 pagesChương 5 - Môn thiết kế và phát triển sản phẩm - IUHNam NguyenHoangNo ratings yet
- Nước ép trái cây kết hợp với rau, củ- Môn thiết kế và phát triển sản phẩm- IUHDocument41 pagesNước ép trái cây kết hợp với rau, củ- Môn thiết kế và phát triển sản phẩm- IUHNam NguyenHoangNo ratings yet
- Xây dựng hệ thống Haccp- chuối sấy dẻoDocument16 pagesXây dựng hệ thống Haccp- chuối sấy dẻoNam NguyenHoangNo ratings yet
- Tổng quan về mối liên hệ giữa dinh dưỡng và di truyền học biểu sinh- IUHDocument41 pagesTổng quan về mối liên hệ giữa dinh dưỡng và di truyền học biểu sinh- IUHNam NguyenHoangNo ratings yet
- Đề Thi Cuối Kì Môn Dinh Dưỡng Lâm Sàng- IUHDocument1 pageĐề Thi Cuối Kì Môn Dinh Dưỡng Lâm Sàng- IUHNam NguyenHoangNo ratings yet
- Chương 2- Môn thiết kế và phát triển sản phẩm - IUHDocument16 pagesChương 2- Môn thiết kế và phát triển sản phẩm - IUHNam NguyenHoangNo ratings yet
- PPT thuyết trình môn chế biến thực dưỡng - IUHDocument29 pagesPPT thuyết trình môn chế biến thực dưỡng - IUHNam NguyenHoangNo ratings yet
- Xác định độ tinh khiết tinh dầu- Môn hóa học hợp chất tự nhiênDocument9 pagesXác định độ tinh khiết tinh dầu- Môn hóa học hợp chất tự nhiênNam NguyenHoangNo ratings yet
- Chương 4 - Môn thiết kế và phát triển sản phẩm - IUHDocument16 pagesChương 4 - Môn thiết kế và phát triển sản phẩm - IUHNam NguyenHoangNo ratings yet
- Quản lý cân nặng cho bệnh nhân thừa thừa - béo phì, suy dinh dưỡng- Môn dinh dưỡng lâm sàngDocument35 pagesQuản lý cân nặng cho bệnh nhân thừa thừa - béo phì, suy dinh dưỡng- Môn dinh dưỡng lâm sàngNam NguyenHoangNo ratings yet
- Béo Phì Ngư I L N - Môn Dinh Dư NG Lâm SàngDocument25 pagesBéo Phì Ngư I L N - Môn Dinh Dư NG Lâm SàngNam NguyenHoangNo ratings yet
- XƯƠNG - THẤP KHỚP N9Document38 pagesXƯƠNG - THẤP KHỚP N9Nam NguyenHoangNo ratings yet
- Chương 1 - Môn thiết kế và phát triển sản phẩm - IUHDocument22 pagesChương 1 - Môn thiết kế và phát triển sản phẩm - IUHNam NguyenHoangNo ratings yet
- Bệnh rối loạn tiêu hóa - word - Môn dinh dưỡng lâm sàngDocument33 pagesBệnh rối loạn tiêu hóa - word - Môn dinh dưỡng lâm sàngNam NguyenHoangNo ratings yet
- Bệnh thận và dinh dưỡng điều trị - Môn dinh dưỡng lâm sàngDocument27 pagesBệnh thận và dinh dưỡng điều trị - Môn dinh dưỡng lâm sàngNam NguyenHoangNo ratings yet
- Bệnh tim mạch- Môn dinh dưỡng lâm sàngDocument43 pagesBệnh tim mạch- Môn dinh dưỡng lâm sàngNam NguyenHoangNo ratings yet
- Bỏng và phẫu - Phòng ngừa và điều trị ung thư- Môn dinh dưỡng lâm sàngDocument38 pagesBỏng và phẫu - Phòng ngừa và điều trị ung thư- Môn dinh dưỡng lâm sàngNam NguyenHoangNo ratings yet
- Danh Sách Thực Phẩm Lên Men- wikivi.icuDocument24 pagesDanh Sách Thực Phẩm Lên Men- wikivi.icuNam NguyenHoangNo ratings yet
- Liệu pháp dinh dưỡng cho đối tượng bị rối loạn gan mật - Dinh dưỡng lâm sàngDocument48 pagesLiệu pháp dinh dưỡng cho đối tượng bị rối loạn gan mật - Dinh dưỡng lâm sàngNam NguyenHoangNo ratings yet
- Đại cương môn tin học - IUHDocument863 pagesĐại cương môn tin học - IUHNam NguyenHoangNo ratings yet
- Thiết kế mô hình kinh doanh các loại bánh quy yến và trà nhằm hạn chế bệnh tiểu đường- Môn quản lý dịch vụ ẩm thựcDocument59 pagesThiết kế mô hình kinh doanh các loại bánh quy yến và trà nhằm hạn chế bệnh tiểu đường- Môn quản lý dịch vụ ẩm thựcNam NguyenHoangNo ratings yet
- Bệnh rối loạn tiêu hóa - ppt- Môn dinh dưỡng lâm sàngDocument42 pagesBệnh rối loạn tiêu hóa - ppt- Môn dinh dưỡng lâm sàngNam NguyenHoangNo ratings yet