Professional Documents
Culture Documents
Ôn tập tế bào học 1
Uploaded by
Alida KimOriginal Title
Copyright
Available Formats
Share this document
Did you find this document useful?
Is this content inappropriate?
Report this DocumentCopyright:
Available Formats
Ôn tập tế bào học 1
Uploaded by
Alida KimCopyright:
Available Formats
Nguyễn Minh Khôi
Các sự kiện trọng đại của sinh học tế bào
- 1655, lần đầu tiên Robert Hooke đưa ra mô tả về tế bào cây bần.
- 1674, 1683, Leeuwenhoek quan sát động vật nguyên sinh và vi khuẩn.
….
- 1838, Schleiden và Schwann giới thiệu thuyết tế bào Cell Theory. 1855 Nageli và Virchow bổ sung thêm
ba nguyên tắc nữa cho học thuyết này.
- 1857 - Kolliker mô tả ty thể.
- 1869 - Miescher lần đầu tiên phân lập DNA .
- 1882 - Kock nhận dạng vi khuẩn.
- 1898 - Golgi khám phá ra phức hệ Golgi.
- 1931 - Ruska tạo ra kính hiển vi điển tử truyền (TEM) đầu tiên.
- 1953 - Watson and Crick đề xuất cấu trúc xoắn kép của DNA.
- 1965 – Chiếc kính hiển vi điện tử quét (SEM) đầu tiên được sản xuất.
Thuyết tế bào
- Tất cả các cơ thể đều chứa một hoặc nhiều hơn một tế bào ( bởi Schwann 1839)
- Tế bào là đơn vị cơ bản của sự sống.
- Tất cả các tế bào phát sinh từ các tế bào tồn tại trước đó (bởi Virchow 1855)
Học thuyết tế bào hiện đại bao gồm một số thông tin thêm:
- Dòng năng lượng diễn ra trong các tế bào.
- Thông tin di truyền (DNA) được truyền từ tế bào này sang tế bào khác.
- Tất cả các tế bào có thành phần cấu tạo hóa học về cơ bản giống nhau.
Các đặc tính của các tế bào
- Các tế bào có tính tổ chức và tính phức tạp cao.
- Các tế bào có một bộ máy di truyền và các phương tiện để sử dụng nó.
+ Lưu trữ thông tin di truyền: tất cả các tế bào - từ vi khuẩn cho tới con người – đều có DNA như
là một dạng lưu trữ thông tin tế bào.
+ Sự biến đổi thông tin: Các thông tin được lưu trữ trong DNA được sao lại thành RNA.
+ Thông tin được sử dụng như thế nào: RNA được sao từ DNA sau đó được dịch thành các protein
bởi tế bào cho các hoạt động của nó.
- Các tế bào có khả năng tự nhân đôi
- Các tế bào thu hái và sử dụng năng lượng.
- Các tế bào thực hiện vô số các phản ứng hóa học.
- Các tế bào tham gia vào nhiều hoạt động cơ học,vận động (Sự chuyển động của tế bào, sự co cơ)
- Các tế bào có thể đáp ứng lại các kích thích (miễn dịch hay tín hiệu tế bào)
- Các tế bào có khả năng tự điều chỉnh (sự phân chia và sự chết tế bào)
Ôn tập Tế bào học pg. 1
Nguyễn Minh Khôi
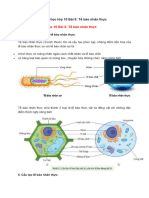
Khái quát các phần của tế bào
- Tế bào chất: chứa các enzym cho đường phân ( một giai đoạn của hô hấp hiếu khí) và các phản ứng chuyển
hóa khác với đường, muối, aminoaxit, nucleotide, và tất cả những gì khác cần để tế bào thực hiện chức
năng.
- Nhân: là bào quan lớn nhất, bao quanh bởi màng nhân là một màng kép với các lỗ nhân; Có chứa nhân
con ( hạch nhân ). Tham gia sản xuất các ribosome.
- Ty thể: là một bào quan dạng cái xúc xích ( thật đấy ) và là vùng hô hấp hiếu khí.
- Lục lạp: là vùng xảy ra quang hợp, giống như ty thể, chúng được bao bởi một màng kép, song chúng cũng
có một màng thứ ba là màng Thilakoid . (Chỉ có ở thực vật)
- Các Ribosome: nơi tổng hợp protein, bao gồm hai tiểu phần, một lớn một nhỏ. Nó được tạo ra từ RNA và
protein.
- Lưới nội chất trơn (SER): một chuỗi các kênh có màng bao bọc, liên quan tới tổng hợp và vận chuyển các
vật liệu, chủ yếu là các lipid cần cho tế bào.
- Lưới nội chất hạt (RER): tương tự lưới nội chất trơn, nhưng được khảm nhiều ribosome tạo ra bề mặt sần
sùi của nó.
- Phức hệ Golgi: một loạt các túi dẹt có màng khác, được hình thành từ lưới nội chất. Nhiệm vụ của nó là
chuyển protein từ RER ra màng tế bào để xuất ra.
- Không bào: là một bào quan có màng bao bọc xuất hiện ở tất cả các tế bào thực vật, nấm và ở một số sinh
vật đơn bào, động vật và vi khuẩn. Chức năng và tầm quan trọng của không bào cũng phụ thuộc đa dạng
vào các dạng tế bào có sự xuất hiện của nó.
- Lysosome: là một bào quan có màng chứa các enzyme hydrolase thủy phân axit để phá hủy các vật liệu
thừa thãi và các mảnh vụn tế bào. Chúng được tạo ra khi thêm các enzyme thủy phân vào các endosomes
mới từ phức hệ Golgi.
- Bộ khung xương tế bào: đây là một mạng lưới các protein sợi trải khắp các tế bào nhân thực, được dùng
để nâng đỡ, vận chuyển và chuyển động.
- Trung thể: là một cặp vi ống ngắn có liên quan tới phân chia tế bào. Trước khi mỗi tế bào phân chia, hai
trung thể di chuyển về hai đầu đối diện của tế bào, nơi mà chúng sử dụng các thoi tơ để tổ chức và tách các
nhiễm sắc thể. (không có ở thực vật và một số động vật)
- Lông và roi: bao phủ trên các lớp niêm mạc.
- Màng sinh chất: là một lớp kép mỏng và dẻo bao quanh bên ngoài tất cả các tế bào, được tạo ra từ
phospholipid và protein. Chúng phân chia các thành phần bên trong tế bào với môi trường và điều khiển sự
ra vào của các vật liệu.
- Thành tế bào: một màng dày hơn màng sinh chất, được dùng để tạo ra sự bền chắc cho tế bào (thực vật).
MÀNG TẾ BÀO
Chức năng cơ bản của màng sinh chất
- Bảo vệ
- Điều khiểu quá trình vận chuyển vật chất ra vào tế bào (thấm chọn lọc)
- Đặc trưng hóa tế bào
- Nâng đỡ và tạo hình tế bào (gắn kết với hệ thống khung xương tế bào)
Mô hình cấu trúc màng sinh chất – mô hình khảm lỏng
- Lỏng (fruid) – mỗi phân tử phospholipid và protein có thể di chuyển từ vị trí này sang vị trí khác trong
cùng một lớp lipid giống như dung dịch.
- Khảm (mosaic) – các phân tử protein nằm xen kẽ vào lớp lipid khi quan sát màng từ trên xuống.
Ôn tập Tế bào học pg. 2
Nguyễn Minh Khôi
*) Phospholipid
- Có đầu phân cực và có tính chất ưa nước (hydrophilic)
- Có đầu không phân cực có tính chất kị nước (hydrophobic)
- Các loại phân tử lipid khác nhau của màng do đó đều lưỡng tính (phosphatidylserine, cholesteron,
galactocerebroside thuộc phospholipid, sterol, glycolipid)
- Các phân tử lipid của màng tế bào xếp thành lớp kép trong nước
+ Màng được tạo thành từ hai lớp phospholipid – gọi là lớp lipid kép
+ Các phospholipid tinh khiết có thể khép kín lại, hạt cầu lipid
- Lớp lipid kép có tính chất lỏng
+ Các phân tử phospholipid có thể di chuyển
+ Có thể chuyển trạng thái lỏng/nhớt: thành phần càng nhiều lipid chưa bão hòa thì càng lỏng, càng
nhiều lipid bão hòa thì càng nhớt.
+ Màng tế bào của động vật có các phân tử Cholesteron (sterol), chúng làm cứng màng tế bào (giảm
tính lỏng)
- Lớp lipid kép là chất lỏng hai chiều : các phân tử lipid trong lớp lipid kép có thể :
+ Chuyển động qua lại xung quanh vị trí của nó theo chiều ngang
+ Quay tròn tại vị trí của nó
+ Đổi chỗ với các lipid bên cạnh
+ Flip – flop tức là phân tử quay đầu, từ lớp này sang lớp khác, hiếm khi xảy ra.
- Lớp kép lipid có tính chất bất đối xứng. Các phân tử phospholipid và các phân tử glycolipid phân bố bất
đối xứng trên màng kép. Flippase, Scramblase tham gia vào quá trình tổng hợp lớp lipid kép (trong đó
scramblase thêm một cách ngẫu nhiên vào một nửa lớp kép, còn flippase thêm một cách định hướng)
*) Protein màng
- Protein màng thực hiện nhiều chức năng: vận chuyển, hoạt tính enzym, thụ thể bề mặt tế bào, dấu ấn của
tế bào, liên kết tế bào, gắn kết màng với bộ khung xương tế bào.
- Các protein màng chia hai loại theo vị trí của nó: nội màng và ngoại màng
Ôn tập Tế bào học pg. 3
Nguyễn Minh Khôi
- Các protein màng liên kết với lớp lipid kép bằng nhiều cách
+ Xuyên màng (transmembrane) một hoặc nhiều lần. Mỗi chuỗi polipeptide thường xuyên màng
với cấu trúc xoắn α
+ Bám vào một mặt của màng kép
+ Thông qua một số trung gian liên kết từ màng với đầu NH2 hoặc COOH
+ Protein đính kèm: đi liền với một protein xuyên màng khác.
- Màng sinh chất được củng cố bằng lớp vỏ của tế bào (cell cortex): hầu hết màng tế bào được nâng đỡ và
hỗ trợ bởi một khung protein được đính vào màng bởi các protein xuyên màng để làm tăng độ bền vững
của màng
- Tế bào hạn chế sự di chuyển của các protein màng: tế bào có một số cách để giam giữ các protein ở các
vị trí xác định trên màng kép bằng cách tạo ra membrane domains trên bề mặt tế bào hay bào quan. Các
protein màng có thể bị buộc với:
+ Các phân tử trong chất nền ngoại bào
+ Một tế bào gần kề
+ Các cấu trúc bất động bên trong tế bào
+ Đặc biệt là với lớp vỏ tế bào
*) Ngoài ra bề mặt tế bào được phủ bởi lớp cacbohydrate (gọi là glycocalyx): lớp cacbohydrate được tạo
thành bởi các chuỗi bên olgosaccharide gắn vào các phân tử glycolipid và glycoprotein của màng.
VẬN CHUYỂN CÁC CHẤT QUA MÀNG SINH CHẤT
- Nồng độ các ion ở phía trong và ngoài màng là rất khác nhau. Ta quan tâm tới nồng độ của Na+ và Ca2+
(ngoài cao hơn trong), của Ka+ (ngoài nhỏ hơn trong)
- Lớp lipid kép ngăn cản các chất hòa tan và các ion:
+ Các phân tử nhỏ, không phân cực (O2, CO2, N2 và các hormone steroid) thấm qua hoàn toàn
+ Các phân tử phân cực nhưng trung hòa điện tích, có kích cỡ nhỏ mới có thể đi qua (H2O, ethanol),
còn kích cỡ lớn (vd như glycerol) không qua được
+ Các phân tử có điện tích bao gồm các ion vô cơ kể cả là chúng có nhỏ thế nào đều bị ngăn lại
- Các protein vận chuyển được chia làm hai loại, khác nhau ở cách mà chúng phân biệt các loại chất tan
khác nhau.
- Mỗi loại màng sử dụng một bộ protein vận chuyển riêng
a) Vận chuyển bị động
*) Khuếch tán đơn giản
- Không đòi hỏi năng lượng
- Theo chiều gradient nồng độ
- VD: O2 và H2O khuếch tán vào và CO2 khuếch tán ra.
*) Khuếch tán có điều kiện
- Không đòi hỏi năng lượng
- Sử dụng các protein vận chuyển để vận chuyển các chất theo chiều gradient nồng độ.
- VD: glucoso và các axitamin vận chuyển từ máu vào tế bào
*) Thẩm thấu
- Do sự khuếch tán của nước qua màng: di chuyển từ nơi có thế nước cao (nồng độ chất tan thấp) đến nơi
có thế nước thấp (nồng độ chất tan cao)
- Kênh vận chuyển nước ( Aquaporin) được sử dụng trong quá trình thẩm thấu.
- Sinh ra môi trường: đẳng trương, ưu trương (nồng độ chất tan lớn hơn nồng độ trong nội bào), môi trường
nhược trương (nồng độ chất tan nhỏ hơn môi trường nội bào)
Ôn tập Tế bào học pg. 4
Nguyễn Minh Khôi
- Nồng độ chất tan bên trong tế bào thường lớn hơn dung dịch ngoại bào (theo lẽ thường nước sẽ đi vào tế
bào làm nó trương lên). Tế bào sử dung một số cách để tránh bị li giải do áp suất thẩm thấu
+ Động vật nguyên sinh thải nước định kỳ ra khỏi tế bào và tích nước trong không bào
+ Thực vật có thành tế bào bảo vệ.
b) Vận chuyển chủ động
- Đặc điểm:
+ Đòi hỏi năng lượng
+ Vận chuyển ngược chiều gradient
+ Dựa vào các protein vận chuyển, điều khiển bởi gradient nồng độ và điện thế màng.
- Có ba dạng vận chuyển : đơn vận, đồng vận, đối vận.
- Các protein vận chuyển : được dùng để vận chuyển hầu hết các chất hữu cơ qua màng. Có tính chất chọn
lọc, thường mỗi protein chỉ vận chuyển một loại phân tử. Mỗi màng chứa một bộ protein màng đặc trưng.
+ Các protein vận chuyển các chất không tích điện theo chiều gradient nồng độ (thụ động) (không
tính đến điện thế vì chúng không tích điện) như glucoso
+ Các protein vận chuyển các chất tích điện (ion) dựa vào gradient điện hóa (nồng độ và điện thế)
(chủ động). Gradient của nồng độ và điện thế có thể cùng chiều hay ngược chiều nhau.
- Vận chuyển chủ động ngược chiều gradient điện hóa.
- Có 3 cách vận chuyển chủ động: vận chuyển nhờ năng lượng ATP vận hành các bơm, vận chuyển cặp
đôi, và vận chuyển nhờ quang năng.
+ Các tế bào động vật sử dụng năng lượng từ quá trình thủy phân ATP để bơm Na+ ra và bơm K+
vào.
Ôn tập Tế bào học pg. 5
Nguyễn Minh Khôi
Các bước vận hành kênh Na – K
+ 3 ion Na+ nội bào bám vào protein làm
protein biến đổi cấu hình.
+ ATP bám vào protein và phosphoryl hóa
nó; protein được thêm 1 Pi và nhả ADP ra.
+ Khi có Pi, protein lại biến đổi cấu hình và
nhả 3 Na+ ra môi trường ngoại bào
+ 2 K+ ngoại bào bám ngay vào protein do
protein đang có ái lực cao với K+.
+ Khi gắn K+, protein lại biến đổi cấu hình
và tách Pi ra
+ Mất Pi, protein quay về cấu hình ban đầu
như khi chưa liên kết với Na+, dạng này ái
lực thấp với K+ nên K+ được nhả vào tế bào.
Một thế gradient nồng độ Na+ được tạo ra giữa màng sinh chất. Bằng các bơm như vậy, nồng độ Na+ tế
bào chất thấp hơn ngoại bào là 10 – 30 lần
Thế gradient nồng độ Na+ hoạt động cùng với gradient điện thế màng
+ Vận chuyển cặp đôi (coupled transporters): các tế bào động vật sử dụng gradient điện hóa Na+ để
vận chuyển glucoso vào tế bào một cách chủ động . Đây là dạng vận chuyển đồng vận. Nhờ có gradient
điện hóa xuyên màng của Na+ mà Na+ và glucoso cùng đi vào các kênh đặc hiệu, tuy nhiên glucoso đang
đi ngược chiều gradient nồng độ của nó
Glucoso có thể được vận chuyển một cách chủ động hay bị động (theo các cách khác nhau) tùy
thuộc vào tình trạng tế bào
Ôn tập Tế bào học pg. 6
Nguyễn Minh Khôi
GLUCOSO được thu nhận
một cách chủ động
Một cách tương tự, các tế bào thực vật cũng thực hiện điều này nhưng thay gradient Na+ bằng
gradient H+
+ Vận chuyển nhờ quang năng tạm thời chưa được nhắc đến ở đây
Các kênh ion và điện thế màng
Ôn tập Tế bào học pg. 7
Nguyễn Minh Khôi
- Các kệnh vận chuyển ion được đặc trưng bởi:
+ Sự chọn lọc các ion
+ Tín hiệu mở kênh
- Các kênh chọn lọc các ion dựa trên 3 yếu tố :
+ Kích thước
+ Hình dạng
+ Điện tích
- Điện thế màng được điều chỉnh bởi sự vận chuyển các ion qua màng
- Các kênh ion được phân loại phụ thuộc vào tín hiệu mở kênh (đặc trưng 2)
- Các kênh ion chuyển đổi trạng thái đóng mở một cách ngẫu nhiên chứ không liên tục duy trì một trạng
thái
Hơn một triệu ion có thể đi qua một kênh ion mỗi giây, tức là nhanh gấp 1000 lần so với tốc độ
của bất kỳ transporter (protein vận chuyển).
c) Các hình thức vận chuyển qua màng khác
- Xuất bào
- Nhập bào
- Uống bào
- Thực bào
PHÂN LOẠI VÀ VẬN CHUYỂN PROTEIN
- Trình tự phân loại hướng dẫn các protein đi đến những nơi mà nó cần thực hiện chức năng.
- Trình tự phân loại thường bị bỏ đi sau khi protein đã được phân loại xong.
Vận chuyển protein vào nhân
*) Lỗ màng nhân
- Gồm hai thành phần : màng trong (các protein làm chỗ bám cho các NST và các sợi protein đan thành
tấm lamina), màng ngoài nối liền với màng lưới nội chất.
- Trên màng nhân có rất nhiều lỗ nhân.
Ôn tập Tế bào học pg. 8
Nguyễn Minh Khôi
- Lỗ màng nhân ngăn cản các phân tử lớn nhưng cho phép nước, và các phân tử tan được trong nước đi
qua.
- Các protein của nhân mới được tổng hợp được chuyển chủ động vào nhân qua lỗ nhân.
- Các protein này mang theo một tín hiệu định vị nhân (nuclear localization signal).
+ Chúng tương tác với các thụ thể (nuclear transport receptors)
+ Các protein nhân được chuyển chủ động bằng năng lượng của GTP
+ Các thụ thể vận chuyển protein sau đó lại được trả lại tế bào chất qua lỗ nhân để tái sử dụng.
+ Các protein được vận chuyển vào với cấu hình không bị biến đổi
Vận chuyển protein qua màng ty thể hay lục lạp.
- Các protein này thường có trình tự phân loại nằm ở đầu N cho phép chúng được chuyển vào 2 bào quan
này
- Chúng được chuyển một cách đồng thời qua cả hai màng tại các vị trí đặc biệt nơi mà hai màng tiếp xúc
với nhau.
- Mỗi protein bị mở ra (mất cấu trúc xoắn ban đầu) khi chúng được chuyển và trình tự tín hiệu bị cắt đi sau
khi quá trình hoàn tất.
- Các protein Chaperone giúp kéo các protein xuyên qua hai màng và giúp tái định hình cấu hình các
protein khi chúng đã vào được bên trong
Ôn tập Tế bào học pg. 9
Nguyễn Minh Khôi
Vận chuyển protein vào trong lưới nội chất
- Lưới nội chất là một hệ thống bể chứa có màng bao bọc (có dạng túi) được giữ với nhau bởi bộ khung
xương tế bào
- Khoang của các bể này nối liền liên tục khắp không gian xung quanh nhân nhưng riêng rẽ với tế bào chất.
- Mạng lưới nội chất cung cấp như một điểm đi vào cho các protein đã được xác định trước cho các bào
quan: Golgi, endosomes, lysosomes, bề mặt tế bào, cũng như chính nó
- Có hai loại protein có thể đi vào lưới nội chất từ chất tế bào: (1) các protein tan trong nước , (2) các protein
xuyên màng tiềm năng.
- Tất cả các protein này trước tiên được dẫn vào lưới nội chất bằng một trình tự tín hiệu, một đoạn gồm 8
aminoaxit kị nước hay nhiều hơn cũng được đòi hỏi đối với quá trình vận chuyển xuyên màng này,
- Hầu hết các protein đi qua màng lưới nội chất trước khi chuỗi polypeptide được tổng hợp hoàn thành
- Yêu cầu các ribosome bám vào lưới nội chất nên mới có lưới nội chất hạt.
Ôn tập Tế bào học pg. 10
Nguyễn Minh Khôi
Ôn tập Tế bào học pg. 11
Nguyễn Minh Khôi
- Tín hiệu khởi đầu và kết thúc xác định sự xắp xếp của protein xuyên màng trong lớp kép.
Ôn tập Tế bào học pg. 12
Nguyễn Minh Khôi
- Hầu hết các protein được thay đổi liên kết cộng hóa trị trong lưới nội chất
+ Cầu lưu huỳnh S-S được hình thành làm bền vững cấu trúc protein
+ Sự glycosyl hóa: oligosaccharide ban đầu đang được gắn vào một lipid đặc hiệu được gọi là
dilichol trong màng lưới nội chất; nó sau đó được chuyển sang vùng có asparagin trong chuỗi protein.
Ôn tập Tế bào học pg. 13
Nguyễn Minh Khôi
Vận chuyển bằng không bào
Ôn tập Tế bào học pg. 14
Nguyễn Minh Khôi
- Cấu trúc Clathrin
*) Rab protein, tethering protein, và SNARES giúp hướng các bóng vận chuyển tới các màng đích của
nó
- Protein tethering gắn trên màng đích sẽ là yếu tố tiếp nhận ban đầu cho các bóng không bào bằng cách
tương tác với protein Rab trên màng của bóng. Từ đó kéo hai màng lại đủ gần nhau
- Sau đó v-SNARE trên bóng (vesicle) sẽ tương tác với t-SNARE trên màng đích (target) để kéo hai màng
gần nhau hơn nữa (~ 1.5 nm)
- Để có được khoảng cách gần như vậy (vốn bị cản trở bởi phân tử nước giữa hai bề mặt màng ưa nước):
ngay khi quá trình dung hợp hai màng bắt đầu, các SNARE quấn lấy nhau và hoạt động như cái máy tời
tạo lực kéo đồng thời đẩy các phân tử nước khỏi vùng tiếp xúc
- Hầu hết các protein đi vào lưới nội chất được xác định cho các vị trí khác. Chúng được đóng gói thành
các túi không bào vận chuyển mọc ra từ lưới nội chất
- Tương tác với chaperon giữ cho các protein trong lưới nội chất cho tới khi việc cuộn gập cẩn thận được
xảy ra, nếu không cuối cùng các protein này sẽ bị thoái biến
Bộ máy Golgi
- được tạo nên từ các chồng của các cấu trúc có màng như các bể
- Mỗi bể chứa có các enzym đặc hiệu của Golgi, chỉnh sửa hay giúp chỉnh sửa các protein hàng hóa đi qua
nó.
- Một số phần thực hiện chức năng: mạng lưới cis – Golgi, trung tâm, mạng lưới trans – Golgi
- Việc chỉnh sửa, phân loại và đóng gói các đại phân tử lớn cho sự tiết tế bào hoặc dùng trong tế bào.
Lộ trình nhập bào
- Quá trình mà tế bào thu nhận các chất lỏng hay các phân tử lớn nhỏ được gọi là thực ẩm bào
(endocytosis)
- Pinocytosis: “Uống bào” (hay ẩm bào), liên quan đến sự ăn các phân tử chất lỏng thông qua các túi tiết
đường kính dưới 150 nm – phổ biến nhất của nhập bào
+ Uống bào thường được thực hiện chủ yếu bởi áo clathrin và các bóng không bào
- Phagocytosis: “Thực bào”, liên quan tới sự ăn các phân tử lớn, như các bào quan nhỏ hay các mảnh vụn
thông qua các túi gọi là phagosome đường kính chừng trên 250 nm.
- Receptor – mediated endocytosis cung cấp một con đường đặc biệt vào tế bào động vật. Quá trình mà
các đại phân tử bám vào các thụ thể trên bề mặt tế bào và đi vào tế bào như những phức hệ thụ thể của đại
phân tử trong túi không bào có áo clathrin
HÔ HẤP HIẾU KHÍ VÀ TY THỂ
Ôn tập Tế bào học pg. 15
Nguyễn Minh Khôi
Ty thể
- Ty thể thường xuất hiện ở những nơi cần tiêu thụ ATP (như tế bào cơ tim hay đuôi tinh trùng cần rất nhiều
ATP)
- Ty thể tiêu thụ oxi
- Được tìm thấy trong các tế bào hiếu khí ở sinh vật nhân chuẩn.
- Tập trung tại các vùng tế bào có hoạt động trao đổi chất mạnh và cần nhiều ôxy
- Ty thể là một hệ thống gồm nhiều hạt ty thể liên kết thành mạng lưới hơn là các cơ quan riêng lẻ
- Rất đa dạng về hình dạng, kích thước, số lượng
- Thời gian sống của ty thể là 10 - 20 ngày
*) Cấu trúc ty thể
- Màng ngoài
+ Bản chất là lipoprotein
+ Chứa nhiều kênh vận chuyển lớn cho phép các phân tử nhỏ có kích thước dưới 5000 Da đi qua
+ Là nơi chứa các enzym liên quan đến quá trình sinh tổng hợp lipid của ty thể
- Khoảng gian màng:
+ Chứa các enzym có khả năng sử dụng ATP mới tổng hợp để photphoryl hóa các nucleotide khác
+ Tham gia vào quá trình bơm proton H+
+ Chứa các protein tham gia vào quá trình apoptosis
- Màng trong:
+ Chứa nhiều protein hơn màng ngoài
+ Cuộn thành các mào răng lược ăn sâu vào chất nền
+ Chứa hệ thống protein tổng hợp ATP
+ Chứa các protein tham gia vào quá trình truyền điện tử
+ Chứa các protein vận chuyển chủ động các chất vào trong chất nền
+ Chứa các kênh vận chuyển ion.
- Chất nền:
+ Chứa hệ enzym cho chu trình Krebs
+ Chứa hệ enzym tổng hợp các axit béo
+ Chứa DNA ty thể, ribosome, tRNA, mRNA, rRNA và các enzym cần cho sự biểu hiện gen
Ôn tập Tế bào học pg. 16
Nguyễn Minh Khôi
*) Phát sinh ty thể
- Chứa DNA ty thể: DNA trần, dạng vòng
- DNA ty thể có khả năng tự tái bản
- Chứa tRNA, mRNA, rRNA
- Tự tổng hợp một số loại protein. Axit amin khởi động là N-formyl-methionine
- Giả thuyêt “cộng sinh”: sự xuất hiện của ty thể là do một loại vi khuẩn hiếu khí cộng sinh vào tế bào
*) Chức năng ty thể
- Là nhà máy sản sinh ATP
- Tham gia các quá trình trao đổi chất
- Tham gia vào quá trình apoptosis
- Tự tổng hợp protein riêng cho ty thể
Sản xuất ATP
*) Giai đoạn 1: Đường phân
- Đường phân sản xuất ra ATP không yêu cầu oxi
- Diễn ra trong tế bào chất
- Đường phân có ở hầu hết các tế bào kể cả các vi sinh vật kỵ khí
- Đường phân là một quá trình gồm một chuỗi 10 phản ứng liên tiếp
+ Bước 1 và bước 3 sử dụng năng lượng thủy phân ATP
+ Bước 4 cắt 1 phân tử đường 6C thành 2 phân tử đường 3C là đồng phân của nhau, trong đó một
đồng phân được đưa đến bước 6, một được đưa đến bước 5 đồng phân hóa rồi mới sang bước 6
+ Bước 6 bắt đầu pha sinh năng lượng (nhưng ATP chưa được sinh từ bước 6)
+ Quá trình hình thành ATP diễn ra ở bước 7 và 10 là sự phosphoryl hóa mức cơ chất – là quá trình
chuyển gốc Pi từ một phân tử cơ chất vào ADP.
+ Năng lượng sinh ra từ đường phân nằm trong hai dạng: ATP và các e cao năng trong NADH.
+ Trong đường phân có phản ứng oxi hóa ở bước 6 mặc dù không có mặt của khí oxi.
C6H12O6 + 2NAD+ + 2ADP + 2Pi 2C3H4O3 + 2NADH + 2H+ + 2ATP + 2H2O
Lưu ý là NADH là một chất mang hoạt hóa chứa các e cao năng
*) Bước ngoặt trong đường phân – sự lên men
- Đối với vi sinh vật kị khí hay một số tế bào động vật thường xuyên hoạt động trong môi trường thiếu oxi
(tế bào cơ chẳng hạn), năng lượng chủ yếu đến từ đường phân.
- Sự lên men: pyruvate của đường phân được biến đổi thành các sản phẩm thải khỏi tế bào:
+ Lactacte trong các tế bào cơ (gây ngộ độc cơ tức sự mỏi cơ)
+ Ethanol, CO2 trong tế bào nấm men
Ôn tập Tế bào học pg. 17
Nguyễn Minh Khôi
Trong khi NADH chuyển các e của nó vào tế bào chất rồi được chuyển hóa ngược thành NAD+
(chứ không đi tiếp vào chu trình Crebs)
*) Nói về hô hấp kị khí (anaerobic respiration)
- Vi khuẩn và sinh vật cổ cũng sinh ra ATP trong điều kiện không có oxi.
- Hô hấp kị khí cũng cần chuỗi truyền điện tử trong khi lên men không có giai đoạn này
- Chất nhận e cuối cùng trong hô hấp kị khí không phải là oxi – đây là điểm khác với hô hấp hiếu khí với
chất nhận e cuối cùng là oxi.
- Chuỗi truyền e của hô hấp kị khí cũng diễn ra ở màng, nhưng là màng sinh chất của tế bào - chứ không
phải màng trong ty thể.
*) Giai đoạn 2 của hô hấp hiếu khí – oxi hóa pyruvate
- Chu trình Crebs = chu trình axit citric = Tricacboxylic cycle axit = TCA
- Pyruvate từ đường phân được bơm tích cực vào chất nền ty thể.
- Tại ty thể, pyruvate được decarboxyl hóa tạo ra một acetyl CoA và một NADH gián tiếp tạo thành
Pyruvate Phức hệ enzyme Acetyl CoA + CO2
Dehydrogenase pyruvate
- Cùng với pyruvate tạo ra từ đường phân, các axit béo tạo ra từ chất béo hay các axit amin được tạo từ
protein cũng đều tạo được Axetyl CoA trong chất nền ty thể.
Lưu ý sản phẩm có cả 1 NADH trong ty thể.
*) Giai đoạn 3: chu trình Crebs
- Trong các vi khuẩn hiếu khí – không có ty thể - đường phân và sự sản sinh acetyl CoA, cũng như chu
trình axit citric được diễn ra tại tế bào chất.
- Trong chu trình Crebs có tạo ra một phân tử FADH2 từ FAD là một chất mang hoạt hóa tương tự NADH,
mang những e (và hydro) cao năng.
- Trong TCA, NADH được sinh ra ở bước 3, 4, 8. Tạo ra FADH2 tại bước 6. GTP được tạo ra ở bước 5 có
ý nghĩa tạo ra 1 ATP vì nó chuyển Pi cho ADP
Ôn tập Tế bào học pg. 18
Nguyễn Minh Khôi
phương trình tổng quát chu trình Crebs
1 Acetyl CoA + 3 NAD+ + 1 FAD + (GDP + Pi) + 2H2O 2 CO2 + 3 NADH + FADH2 + GTP + CoA
Chú ý: Các phản ứng tạo ra NADH và FADH2 là các phản ứng oxi hóa
*) Giai đoạn cuối: Sự oxi – phosphoryl hóa
- Sự vận chuyển điện tử vận hành quá trình tổng hợp phần lớn năng lượng ATP của hầu hết các tế bào.
- Trong quá trình oxi – phosphoryl hóa, các chất mang NADH, FADH2 truyền các điện tử cao năng cho
chuỗi dẫn truyền điện tử
- Một hệ thống các chất vận chuyển điện tử trong màng trong ty thể thực hiện chuỗi vận chuyển.
- Tại một số vùng đặc biệt của chuỗi, năng lượng đã giải phóng của các e dùng để bơm các H+ từ chất nền
ty thể ra khoảng gian màng
- Các điện tử đạt mức năng lượng thấp nhất
- Sự oxi – phosphoryl hóa xảy ra ở cả các tế bào nhân thực và các vi khuẩn kiếu khí.
- Sự oxi phosphoryl hóa được chia làm hai giai đoạn: vận chuyển điện tử và hóa thẩm thấu (tức tổng hợp
ATP)
*) Chuỗi truyền điện tử.
- Quá trình oxy hóa khử coenzym bằng cách truyền điện tử đến oxi phân tử được gọi là sự truyền điện tử
- Sự truyền điện tử liên quan đến các phản ứng oxi hóa giải phóng năng lượng, trong đó NADH và FADH2
được oxi hóa và oxi là chất nhận e cuối cùng.
- Sự truyền điện tử luôn tiêu thụ O2 và tạo ra H2O
- Sự truyền điện tử liên quan đến một loạt các chất mang điện tử có khả năng oxi háo thuận nghịch. Tập
hợp các chất này được gọi là chuỗi truyền điện tử
- Chuỗi truyền điện tử bao gồm 4 phức hệ chính: phức hệ NADH dehydrogenase; phức hệ succinate
dehydrogenase; phức hệ cytochrome b-c1; phức hệ cytochrome a và a3 (còn gọi là cytochrome
oxidase).
-Các chất mang điện tử hoạt động theo một thứ tự xác định dựa vào thế oxi hóa khử của chúng.
- Các e được chuyển qua ba phức hệ enzym nằm xuyên màng trong ty thể.
Ôn tập Tế bào học pg. 19
Nguyễn Minh Khôi
- Sự di chuyển của các điện tử qua các phức hệ hô hấp này đi kèm với việc bơm các proton từ chất nền ty
thể ra khoảng gian màng mỗi phức hệ enzym là một bơm proton.
- Trong bản chất, các e đi ra từ các chất mang điện tử có ái lực e thấp tới các chất có ái lực e cao hơn cho
tới tận khi gặp O2 là chất nhận e cuối cùng (ái lực lớn nhất) thế oxi hóa khử liên tục tăng trong chuỗi
truyền điện tử và sự truyền e trong chuỗi là một quá trình lợi về năng lượng
- Việc bơm H+ ra ngoài khoảng gian màng tạo ra một gradient điện hóa trên màng trong ty thể.
*) Hóa thẩm thấu – vận hành tổng hợp ATP
- Sự xác lập gradient điện hóa : dựa vào điện thế màng và nồng độ các ion.
+ Đầu tiên, các proton được bơm ra ngoài khoảng gian màng sinh ra một gradient H+ giữa hai phía
của màng trong. Lúc này pH của chất nền cao hơn pH của gian màng là 0.7 đơn vị.
+ Sự bơm proton ra tạo ra một gradient điện thế màng, vùng chất nền sát màng tích điện âm, vùng
gian màng sát màng tích điên dương
Tạo ra một thế rất mạnh kéo H+ quay trở lại chất nền.
- ATP synthase:
+ Là một phức hệ protein gồm phần có hoạt tính enzym nằm quay về phía chất nền ty thể.
+ Enzym này có cấu tạo gồm đầu F1 và phần xuyên màng làm nhiệm vụ dẫn H+ gọi là Fo
- Hoạt động của ATP synthase
Ôn tập Tế bào học pg. 20
Nguyễn Minh Khôi
(Để ý rằng mỗi lần quay một góc 120o tức là F1 đã thay đổi cấu hình)
+ Đầu Fo của ATP synthase có 10 vị trí giống nhau để proton có thể đi qua và tương tác. Mỗi 3
proton đi qua (F1 thay đổi cấu hình 3 lần) thì tạo một ATP.
+ Một vòng quay 360o của Fo tạo ra 3 ATP. Với tốc độ tốt 100 ATP có thể được tạo ra trong 1 giây.
- ATP synthase có thể quay ngược lại như là một bơm proton và cũng sử dụng năng lượng thủy phân ATP.
Tổng kết cả quá trình:Đường phân: 2ATP (bước 7, 10) + 2 NADH (bước 6) = 2ATP + 2x2 = 6ATP
Oxi hóa pyruvate: 2NADH = 2x3 = 6ATP
TCA:2 ATP (từ GTP bước 5)+ 2 FADH2 + 6 NADH = 2ATP + 2x2 + 6x3 = 24ATP
36 ATP ở tế bào nhân thực và 38 ATP ở tế bào nhân sơ (2 NADH tạo ra từ đường phân nằm ngoài ty thể
nên nó trải qua một số lọ trình khác thì mới chuyển các e của nó cho ty thể, do đó chỉ tạo ra 2 ATP/NADH)
BỘ KHUNG XƯƠNG TẾ BÀO
a) Vi ống
- Vi ống là một trong ba cấu trúc chính của bộ khung xương tế bào nhân thực
- Chức năng chính:
+ Các vi ống (MTs) cung cấp lực cơ học để duy trì hình dạng của tế bào
+ Cùng với các protein vận động tham gia vào vận chuyển nội bào
+ Phân tách các nhiễm sắc thể trong quá trình phân chia
*) Cấu trúc và sự hình thành các vi ống.
- Các nhị hợp tubulin là các đơn vị cấu trúc của MTs
Ôn tập Tế bào học pg. 21
Nguyễn Minh Khôi
- Nhị hợp tubulin gồm hai phân tử protein hình cầu: α – tubulin và β – tubulin
- Các MTs là các sợi phân cực : đầu (+) và đầu (-)
- Hai đầu của MTs khác nhau về cấu trúc và tính chất
- Mỗi MT được cấu tạo từ 13 sợi nguyên (protofilament)
- Nhị hợp α – tubulin và β – tubulin trọng lượng phân tử 55 kDa tương đồng về cấu trúc. Mỗi protein chứa
một vị trí liên kết với nucleotide
+ α – tubulin liên kết với GTP, phân tử này không bị thủy phân
+ β – tubulin liên kết với GDP hoặc GTP. Trong một số điều kiện β – tubulin có thể thủy phân
GTP thành GDP + Pi và liên kết với GTP
GTP cần phải được liên kết với cả tiểu đơn vị α β thì các nhị phân mới liên kết được với nhau để hình
thành sợi nguyên và vi ống.
Đầu (-) của vi ống đóng vai trò trong việc hoạt hóa hoạt tính của β – tubulin
*) Sự hình thành MTs
- Các vi ống được hình thành bằng sự thêm các nhị hợp vào hai đầu của chúng
- Các vi ống có thể tạo thành dạng đơn chuỗi, chuỗi đôi hay chuỗi ba
*) Các số liệu
- Đường kính ngoài ống : 25 nm
- Đường kính trong ống : 15 nm
- Đường kính một tubulin đơn : 4 – 5 nm ; khối lượng phân tử 55kDa
- Hai tubulin dính vào nhau dài 8 nm
- Mỗi MT có 13 sợi nguyên
- MTs thẳng, dạng trụ rỗng. Chiều dài thay đổi thường dưới 200 nm, với vi ống sợi trục dài nhiều micromet
*) Treadmilling
- Khi nồng độ tubulin tự do cao hơn nồng độ cân bằng ở đầu (+) nhưng nồng độ tubulin tự do lại thấp hơn
ở đầu (-), sự thêm nhị hợp ở đầu dương đồng thời loại bỏ nhị hợp ở đầu âm với tốc độ ngang nhau.
*) Sự tổ chức MTs trong tế bào
- Trung tâm tổ chức vi ống (MTOC) là nơi sự hình thành vi ống được khởi đầu và đóng vai trò như nơi neo
giữ một đầu của các vi ống này
- Ở tế bào động vật, MTOC là trung thể: gồm các trung tử và vật liệu quanh thể trung tâm (pericentriolar
matrix)
- Các phức hệ γ – tubulin vòng của trung thể đáp ứng quá trình hình thành vi ống mới. Nếu thiếu các protein
này, trung tử không thể tạo ra gốc vi ống.
- Các vi ống có đặc tính dynamic instability
+ MTs phát triển nhanh chóng ở đầu dương (tận cùng là β – tubulin) Và rất chậm ở đầu âm (tận
cùng là (α – tubulin)
+ Đầu dương thường chuyển giữa hai pha: pha phát triển chậm và pha co ngắn nhanh
+ Mỗi vi ống phát triển và co ngắn một cách độc lập với các vi ống khác.
+ Dải vi ống luôn không ngừng thay đổi chính là khi vi ống mới dài ra và vi ống cũ co lại.
+ Sự thủy phân GTP đóng góp vào hiện tượng này: do liên kết GTP-GTP bền hơn GTP-GDP
+ Ở nồng độ tubulin thấp, các vi ống phát triển chậm do GTP thủy phân quá nhanh, quá trình
giải trùng hợp MTs.
* ) Các protein vận động
- Cấu trúc thường hai phần
Ôn tập Tế bào học pg. 22
Nguyễn Minh Khôi
+ Phần đuôi hình sợi và phần đầu hình cầu
+ Phần đuôi liên kết với các chất cần vận chuyển
+ Phần đầu tiếp xúc trên các vi ống và di chuyển trên các vi ống đó. Tại phần đầu có vị trí liên
kết ATP, và nó có khả năng hoạt động như ATPase thủy phân ATP tạo năng lượng làm thay đổi cấu hình
protein tại đầu hình cầu, giúp protein di chuyển
- Phần lớn các linear motor protein là những protein vận chuyển hàng hóa trong tế bào
Kinesin : Các kinesin khác nhau sẽ di chuyển theo chiều dương hoặc chiều âm của vi ống, hầu hết là
theo chiều dương. Tuy nhiên với mỗi loại kynesin nhất định sẽ chỉ di chuyển theo một chiều. Có hai loại
kinesin là cytosolic kinesin và mitotic kinesin phụ thuộc vào hàng hóa mà đuôi nó xác định. Trong khi
không có hàng hóa, kinesin gập lại. Hai đầu hình cầu của kinesin thay nhau bám vào β-tubulin trên MT,
một cái bám chặt MT trong trạng thái đang thủy phân ATP trong khi cái còn lại liên kết với ADP đang bước
lên.
Các chỉ số (Mỗi bước dài: 8 nm; lực sinh ra: 6 pN; tốc độ: 3µm/sec)
Dynein : Là một phức hệ đa phân lớn, di chuyển theo chiều âm của vi ống. Chúng cần dynamtin để
gắn các bóng không bào hoặc các nhiễm sắc thể với chuỗi nhẹ của nó. Dynein tham gia cấu trúc lông, roi
+ Cấu trúc 9+2 của các vi ống trong các lông/roi của tế bào. Ở đây dynein gồm hai cánh tay dài,
ngắn thò ra
Ôn tập Tế bào học pg. 23
Nguyễn Minh Khôi
b) Sợi trung gian Intermediate filaments (IF)
- Cung cấp hệ thống nâng đỡ cho màng sinh chất, nơi mà chúng tiếp xúc với các tế bào khác hay với các
chất nền ngoại bào.
- Sợi trung gian là dai nhất và bền nhất trong các sợi của khung xương tế bào:dù cho xử lý bằng dung dịch
muối đậm đặc hay bằng các chất tẩy rửa không có tính ion, sợi trung gian vẫn tồn tại.
- IFs bền và giống một sợi dây thừng gồm các sợi protein dài, xoắn. Thứ tự hình thành: monomere là protein
gồm đầu COOH và đầu NH2 hai sợi protein xoắn vào nhau thành dimer vẫn gồm đầu C và N 2 dimer
ghép với nhau gần như so le thành tetramere 8 tetramere hợp lại nhiều bộ 8 tứ hợp ghép lại Sợi
trung gian
IFs không có tính phân cực như vi ống
- Các sợi trung gian chia làm 4 nhóm (ba nhóm đầu trong tế bào chất, nhóm thứ 4 trong nhân) có các tiểu
phân protein tương ứng.
+ keratin trong các tế bào biểu mô. Đa dạng nhất
+ vimentin và họ vimentin trong các tế bào mô liên kết, tế bào cơ hay tế bào thần kinh đệm
+ neurofilaments trong các tế bào thần kinh
+ nuclear lamins làm tăng tính bền vững của màng nhân
- Nhiều sợi trung gian còn được làm ổn định và vững vàng hơn bởi các protein phụ như plectin liên kết
ngang các sợi tạo thành các bó.
- Màng nhân được nâng đỡ bởi mạng lưới sợi trung gian: lớp lamina của nhân được hình thành từ một nhóm
protein sợi trung gian lamin (đừng nhầm với laminin)
+ Lamina được làm tan ra mỗi khi tế bào nhân chia do quá trình phosphoryl hóa các protein lamin
(nhờ các protein kinase) và được tái hình thành nhờ quá trình dephosphoryl hóa lamin (nhờ protein
phosphastase ) ở cuối nguyên phân.
c) Vi sợi actin
- Actin là đơn vị cấu trúc của vi sợi
+ Mỗi phân tử actin bao gồm 4 domain
+ ATP liên kết với actin tại khe domain 2 và 4 cùng với Mg2+
Ôn tập Tế bào học pg. 24
Nguyễn Minh Khôi
- Actin có thể thủy phân ATP ADP, mỗi phân tử Actin có thể liên kết với ATP hay ADP. Cấu trúc không
gian của actin sẽ thay đổi tùy vào liên kết với ATP hay ADP
- Có hai loại actin:
+ G – actin (Global – actin): hình cầu khi liên kết với ATP. Có thể trùng hợp thành F – actin.
Mặt khác có thể giải phóng GDP để liên kết ATP (thường có nồng độ cao hơn trong tb)
+ F – actin (Filament – actin) có thể thủy phân ATP thành ADP . ADP không thể giải phóng
khỏi F - actin vì lúc này khe hở bị chặn
- Actin là những sợi mảnh và linh hoạt: đường kính chỉ khoảng 7 nm, thường ngắn hơn các vi ống (do số
lượng lớn nên tổng độ dài vẫn dài hơn tổng vi ống) nhưng dẻo dai hơn vi ống
- Thường được tìm thấy trong các bó và mạng lưới liên kết chéo.
- Sợi actin cũng có tính phân cự: các đơn phân actin sắp xếp theo cách là tất cả các khe hở (giữa hai domain
2,4) đều quay về một đầu là đầu (-). Đầu còn lại là đầu dương
+ Đầu hình cầu của myosin bám vào actin, tạo hình dạng mũi tên khi quan sát dưới kính hiển vi,
đầu mũi tên chỉ về đầu (-)
- Các đơn phân actin cuốn xung quanh trục của sợi tạo nên cấu trúc xoắn kép.
- Vi sợi và vi ống được trùng hợp theo những cách giống nhau
+ Sự cấu tạo hạt nhân (Nucleation) : trimere của actin cộng lại và hình thành vùng cấu trúc hạt
nhân. Quá trình này chậm chễ.
+ Sự kéo dài (Elongation) : quá trình thêm các phân tử actin để hình thành polymere xoắn. Sau
thời kỳ phát triển, một pha cân bằng đạt được, tại điểm này quá trình giải trùng hợp điều khiển độ dài khi
có các monomere mới được thêm vào.
- Vi sợi cũng có hiện tượng treadmilling
*) Nhiều protein gắn với actin và thay đổi tính chất của chúng
5 % tổng protein trong một tế bào động vật là actin, nồng độ đơn phân actin tự do trong tế bào cũng rất cao
- Thymosin và profilin giúp bắt giữ các đơn phân actin trong tế bào chất, ngăn cản chúng thêm vào đầu
sợi một cách tự do. Do đó điều hòa quá trình trùng hợp actin.
- Formin và actin-related proteins (ARPs) thúc đẩy trùng hợp actin.
Ôn tập Tế bào học pg. 25
Nguyễn Minh Khôi
- Protein Actin-bundling giữ các sợi actin cùng trong dạng bó song song trong microvilli (lông nhỏ), một
số khác liên kết bắt chéo giữa các sợi actin với nhau trong một mạng lưới dạng gel bên trong lớp vỏ tế bào
(cell cortex)
- Các protein Filament-severing cắt các sợi actin thành các độ dài ngắn hơn và vì thế có thể chuyển một
actin gel thành dạng lỏng hơn
- Các sợi actin còn liên kết với protein vận chuyển myosin để hình thành các bó co rút (sợi cơ)
- Capping protein bám vào các đầu của vi sợi, chúng có thể làm bền vững hóa vi sợi (Tropomodulin bịt
đầu âm) hay ức chế trùng hợp, thúc đẩy quá trình phân giải (CapZ capping protein bịt đầu dương)
*) Tế bào bò trườn là nhờ các actin
- Để đẩy rìa tế bào nhô về phía trước, diễn ra trùng hợp vi sợi; mép ngoài của tế bào hình thành hai dạng
lamellipodia (chân giả dạng tấm) và filopodia (chân giả dạng sợi), bên trong hình thành contractile
bundle (bó co)
+ Filopodia (tua nhọn) dài và mảnh. Các bó của sợi actin song song và có đầu dương hướng về phía
chóp filopodia, được liên kết ngang bởi một protein nhỏ như fascin. Khoảng cách gần nhau của các actin
tạo ra độ cứng
+ Lamellipodia dạng lá mỏng chứa một tấm lưới dày đặc các sợi actin được định hướng, đầu dương
hướng ra màng sinh chất tức là hướng ra rìa.
+ Tress fibers hình thành khi một tế bào tạo ra sự tiếp xúc chắc chắn với nền. Các bó của sợi actin
chạy từ rìa phái đuôi đến rìa phía trước của tế bào.
- Cả filopodia và lamellipodia đều có cấu trúc thăm dò và động để hình thành và rụt lại với tốc độ tuyệt vời,
chuyển động xung quanh 1µm mỗi giây.
- Quá trình dàn mỏng của lamellipodia có sự trợ giúp của các ARPs, giúp sự hình thành actin ở rìa phía
trước liên tục và phân giải ở rìa phía sau.
- Filopodia phụ thuộc vào formin, đính vào đầu dương của sợi actin và thúc đẩy hình thành dạng thẳng,
không phải các nhánh.
*) Sự co cơ
- Tất cả các motor protein phụ thuộc vào sợi actin đều thuộc họ myosin, myosin I và myosin II là phong
phú nhất trong tế bào.
+ Myosin I có mặt trong tất cả các tế bào
+ Trong khi đó các tế bào cơ sử dụng chuyên biệt myosin II
- Myosin I
Ôn tập Tế bào học pg. 26
Nguyễn Minh Khôi
+ Có vùng đầu và đuôi, dài tổng cộng 70 nm. Đầu bám vào actin và thủy phân ATP
+ Đuôi của các myosin I khác nhau xác định hàng hóa (túi tiết hay màng sinh chất)
+ Đi theo chiều dương của actin
- Myosin II
+ Có cấu trúc nhị phân, phần đầu hình cầu và phần đuôi dài xoắn với nhau.
+ Các phân tử myosin II có thể liên kết lại thành sợi myosin
+ Actin trượt ngược chiều với myosin trong quá trình co cơ
+ Một tơ cơ (myofibril) (đường kính 1 – 2 µm) gồm một chuỗi các đơn vị nhỏ giống nhau là
sarcomeres (dài tầm 2.5 µm). Sự chồng lên nhau của các sarcomeres tạo thành “cơ vân”
+ Sarcomeres là dạng tập hợp actin với myosin II ở mức cao: các myosin (sợi dày) nằm ở giữa mỗi
sarcomere trong khi các actin (sợi mảnh) kéo dài từ giữa ra hai đầu sarcomere nơi (đầu dương của ) chúng
được cố định bởi đĩa Z
Ôn tập Tế bào học pg. 27
Nguyễn Minh Khôi
+ Sự co các tế bào cơ được gây ra bởi sự thu ngắn đồng thời của tất cả các sarcomere của tế bào, do actin
trượt qua các sợi myosin
+ Không có sự thay đổi độ dài của cả hai loại sợi này.
+ Các chỉ số: 1 sợi myosin có ~ 300 đầu myosin; mỗi đầu myosin gắn – tách actin 5 lần/s; tốc độ trượt
lên nhau của 2 sợi: 15 μm/s đủ để làm cho sarcomere từ trạng thái duỗi cực đại (3μm ) tới co cực đại (2μm )
trong vòng chưa đầy một phần mười giây.
+ Sự co cơ được bấm nút khi nồng độ Ca2+ tăng đột biến trong tế bào chất
TÍN HIỆU TẾ BÀO
- Cơ chế dẫn truyền tín hiệu: tín hiệu điện và tín hiệu phân tử
- Các dạng tín hiệu: tín hiệu nội tiết (endocrine signalling), tín hiệu cận tiết (paracrine signaling) trong
đó có một kiểu đặc biệt là tín hiệu tự tiết (autocrine signalling), tín hiệu thần kinh hay tín hiệu synap
(neuronal signalling) và tín hiệu liên hệ phụ thuộc hay tín hiệu trực tiếp (contact-dependent signaling)
Ôn tập Tế bào học pg. 28
Nguyễn Minh Khôi
- Mỗi phối tủ (ligand) bám với một thụ thể đặc hiệu
- Mỗi tế bào sẽ có những thụ thể riêng phụ thuộc vào chức năng của tế bào đó.
- Hầu hết các tín hiệu ngoại bào sẽ gắn với các thụ thể trên màng tế bào.
- Những phân tử ưa nước có thể xuyên qua màng tế bào để bám trực tiếp các thụ thể nội bào hoặc các
enzyme
- Những con đường mà tín hiệu có thể kết hợp:
+ Một thụ thể nhiều lộ trình
+ Hai thụ thể cho một lộ trình
+ Các thụ thể khác nhau thì hoạt hóa các lộ trình khác nhau;mỗi lộ trình lại ảnh hưởng khác nhau.
- Các loại tế bào khác nhau sẽ đáp ứng khác nhau đối với cùng một loại tín hiệu (acetylcholine sẽ làm tăng
nhịp tim khi tiếp nhận bởi các tế bào điều hòa nhịp tim, là co cơ với các tế bào cơ hay làm tiết dịch vị với
các tế bào tuyên nước bọt)
- Tế bào giải mã tín hiệu của mối liên kết đặc hiệu giữa thụ thể-phối tử để thay đổi sự biểu hiện gen hoặc
phản ứng của tế bào.
- Tín hiệu từ một thụ thể bề mặt tế bào sẽ được chuyển cho một bộ các phân tử tín hiệu nội bào và tác động
tới các efector protein rồi cuối cùng đưa đến những thay đổi trong hành vi của tế bào.
thay đổi có tính chất dây truyền
Ôn tập Tế bào học pg. 29
Nguyễn Minh Khôi
- Con đường dẫn truyền tín hiệu có khả năng khuếch đại tín hiệu để tế bào đáp ứng lại với những tín hiệu
ngoại bào
- Phản ứng của một tế bào với một tín hiệu có thể nhanh hoặc chậm
+ Đáp ứng nhanh là do tất cả các thành phần trong lộ trình đều sẵn sàng: từ các tín hiệu nội bào
(chất truyền tin thứ 2) đến các effector protein đều đang trong tình trạng chờ sẵn. Ví dụ tín hiệu acetylcholine
làm co cơ trong vài milli giây hay làm tiết nước bọt trong vài phút.
+ Đáp ứng chậm là do đáp ứng chỉ xuất hiện chừng nào tế bào có thay đổi trong biểu hiện gene (rồi
phiên mã, dịch mã) tạo ra protein…suy cho cùng là khác nhau về lộ trình. VD: tím hiệu tế bào phân chia
và lớn lên sẽ phải tốn nhiều giờ để biểu hiện.
- Một số hormone đi quang màng sinh chất và bám vào thụ thể nội bào
+ Các tín hiệu quá ưa nước hoặc quá lớn sẽ bám vào các thụ thể bề mặt màng tế bào. Nhóm này rất
lớn
+ Các tín hiệu không quá ưa nước (có thể kị nước) hoặc đủ nhỏ sẽ qua màng và bám lên các thụ
thể nội bào từ đó có khả năng thay đổi biểu hiện gene
+ Nhóm hormone kị nước qua được màng và bám vào các protein thụ thể (trong tế bào chất hay
nhân) gồm: hoormone steroid (cortisol, estradiol, testosteron) và các hormone thyroid như là thyroxine
- Một số chất khí hòa tan qua màng sinh chất và kích hoạt một cách trực tiếp các enzyme nội bào.
+ Điều này có thể tạo ra các thay đổi trong tế bào đích trong một vào giây.
+ Thời gian tác động của các chất khí cũng không kéo dài bao lâu ( do nó tan được và có thể tham
gia phản ứng hóa học)
Một ví dụ đáng quan tâm là NO trong tế bào bám vào và hoạt hóa enzyme guanylyl cyclase thúc
đẩy hình thành GMP vòng
- Các thụ thể bề mặt tế bào chuyển tiếp các tín hiệu ngoại bào thông các lộ trình tín hiệu nội bào
+ Tín hiệu không qua màng được phải cần cầu nối là các thụ thể bề mặt màng tế bào
+ Quá trình truyền tín hiệu nội bào như là cuộc chạy tiếp sức. Quy chung, các thành phần trong lộ
trình tín hiệu nội bào có các chức năng: chuyển tiếp tín hiệu đi xa, khuếch đại tín hiệu, hợp nhất tín hiệu từ
các lộ trình, phân phối các tín hiệu đến các effcetor protein
Ôn tập Tế bào học pg. 30
Nguyễn Minh Khôi
- Một số phân tử tín hiệu nội bào hoạt động như là các công tắc phân tử
+ Sự tiếp nhận tín hiệu (từ giai đoạn trước đó) chuyển các protein tín hiệu nội bào từ ức chế sang
kích hoạt rồi kích thích ( hoặc ức chế ) một (số) protein ở giai đoạn sau trong lộ trình
+ Các protein tín đang hoạt động cũng cần có cơ chế tắt chúng đi.
Việc bật tín hiệu lên cũng quan trọng như việc tắt nó đi.
+ Các protein hoạt động kiểu công tắc này hầu hết thuộc hai nhóm:
Bật tắt nhờ phosphoryl/dephosphoryl hóa (thông qua hoạt tính protein
kinase/protein phosphatase). Thậm chí việc điều khiển là do một lớp các enzym
phosphorylation cascade mà ở đây các protein kinase trước hoạt hóa các kinase
sau sau khi đã khuếch đại, tổng hợp và phân phối thông tin. Phổ biến nhất là
serin/threonin kinases và tyrosin kinases
Bật tắt nhờ GTP – binding proteins (thông qua trạng thái đang liên kết với GTP
hay GDP) Khi được liên kết với GTP, GTPase của nó thủy phân GTP này để lấy
năng lượng và hoạt động, đồng thời chuyển sang liên kết với dạng GDP ngay sau
đó bất hoạt tức là chúng tự dừng lại.
- Các thụ thể bề mặt tế bào rơi vào 3 nhóm
+ Ion-channel-coupled receptors thay đổi tính thấm của màng sinh chất
+ G-protein-coupled receptors kích hoạt trimeric GTP-binding proteins (G proteins) trên màng
+ Enzyme-coupled receptors vừa hoạt động như là các enzyme hoặc liên kết với các enzyme trong
tế bào
Ôn tập Tế bào học pg. 31
Nguyễn Minh Khôi
- Chất truyền tin thứ hai: là các chất nhận lại tín hiệu thông qua các thụ thể rồi sau đó kích thích tế bào
đáp ứng tín hiệu ban đầu. Các chất truyền tin thứ hai như: AMP vòng, GMP vòng, phosphoinositol , axit
arachidonic, ion Ca2+
*) G protein và chất truyền tin thứ hai
- G protein có 3 tiểu đơn vị: α, β, γ
+ Tiểu đơn vị α gắn với GTP và có chức năng thủy phân GTP thành GDP + Pi do chứa GTPase.
+ β và γ có liên kết cộng hóa trị bằng các đuôi lipid ngắn tại vị trí G-protein bám lên màng tế bào
- Sự kích thích đối với GPCRs hoạt hóa các đơn vị G-protein
- Trong trạng thái chưa kích thích, tiểu phần α liên kết GDP. Không có điều gì hay ho xảy ra hết
- Khi GPCRs của nó nhận được tín hiệu ngoại bào, GPCRs thay đổi cấu hình dẫn tới tiểu phần α giảm ái
lực với GDP, sau đó được đổi bằng một phân tử GTP <=> G protein được hoạt hóa tiểu phần α và
phức hệ βγ tách nhau ra để tương tác với các protein đích tương ứng trên màng hoặc chuyển tiếp thông
điệp
Ôn tập Tế bào học pg. 32
Nguyễn Minh Khôi
- Một số G protein trực tiếp điều chỉnh các kênh ion
- Nhiều G protein hoạt hóa các enzym bám màng để tạo ra các phân tử truyền tin thứ cấp
Hai enzyme đích thường xuyên nhất của G protein là:
+ Adenyl cyclase, chúng sản xuất một phân tử tín hiệu thứ cấp nội bào là AMP vòng (cyclic
AMP)
+ Phospholipase C sinh ra hai phân tử tín hiệu nội bào thứ cấp là inositol triphosphate (IP3) và
diacylglycerol (DAG) ( lưu ý thêm là IP3 còn thúc đẩy tích tụ một chất truyền tin thứ 2 khác trong tế bào
chất là Ca2+)
*) Về cAMP
- Là chất truyền tin thứ cấp, được tạo ra từ ATP trong tế bào chất nhờ protein adenyl cyclase. Quá trình
này được khởi động bởi một G protein là GS
Ôn tập Tế bào học pg. 33
Nguyễn Minh Khôi
-
- Quá trình ngược lại được xúc tác bởi các enzym cyclic AMP phosphodiesterase
- Đích tác động của cAMP là cyclic AMP-dependent protein kinase (PKA)
- PKA phosphoryl hóa nhiều protein bằng cách chuyển Pi từ ATP vào vào nhóm chức của serine/threonin
trên protein đích
*) Về Ca2+
- Ca2+ là một chất truyền tin thứ 2, được tích tụ nhờ IP3
Ôn tập Tế bào học pg. 34
Nguyễn Minh Khôi
- Trong tế bào chất có nhiều protein đáp ứng Ca2+, trong đó chủ yếu là calmodulin.
- Calmodulin cũng chủ yếu nhắm vào một đối tượng là Ca2+/calmodulin – dempendent protein kinase
(CaM – kinase).
- Khi các kinase được calmodulin hoạt hóa, chúng phosphoryl hóa có chọn lọc trên các protein đích
MÔ
Các đặc điểm sinh học của mô
- Khả năng biệt hóa (Differentiation)
- Khả năng giải biệt hóa (Dedifferentiation)
Ôn tập Tế bào học pg. 35
Nguyễn Minh Khôi
- Tái sinh/sửa chữa (Regeneration)
- Chuyển dạng (Transformation/metaphasia)
- Bảo quản (Preservation)
- Lão hóa và chết (Aging and death)
Mô gồm hai thành phần
- Chất nền ngoại bào
- Liên kết tế bào (cells – cell junction)
Các loại mô
- Mô liên kết
- Mô thần kinh
- Biểu mô
- Mô cơ
*) Mô liên kết
- Mềm và trong ( như thạch, lấp đầy bên trong mắt)
- Hấp thụ chấn động (sụn)
- Cứng, đặc (xương) hay đàn hồi
- Dẻo và dai (gân, da)
- Các tế bào chính thuộc mô liên kết
+ Nguyên bào sợi (fibroblast): tiết ra các sợi và chất gel vô định hình của chất nền.
+ Đại thực bào (Macrophages): các tế bào thực bào phát triển từ bạch cầu đơn nhân (Monocytes)
+ Tương bào (Plasma cell): các tế bào tiết kháng thể phát triển từ các tế bào lympho B
+ Dưỡng bào (Mass cells): tạo ra histamine giúp mở rộng các mạch máu nhỏ trong phản ứng với
chấn thương
+ Tế bào mỡ (Adypocytes): các tế bào chất béo được lưu trữ dạng triglycerides, nâng đỡ , ngăn
cách và bảo vệ
Chất nền ngoại bào
Bao gồm
- Axit Hyaluronic: phức hợp của các polysaccharides và các protein được tìm thấy trong các mô liên kết
điển hình
- Chondroitin sulphat: chất gel như thạch của sụn, xương, da và mạch máu
- Các chất khác:
- Dermatin sulfate, keratin sulfate, và adhesion proteins
Chất nền ngoại bào chịu lực tác động và có vai trò kết nối, lấp đầy khoảng gian bào.
Các loại sợi thuộc về mô liên kết
- Sợi Collagen: sợi lớn, được làm từ nhiều sợi,
- Sợi Elastic (đàn hồi): các sợi trung gian được tạo bởi các protein elastin, sợi phân nhánh, có thể đàn hồi.
- Sợi Reticular (sợi lưới) : sợi mảnh phân nhánh có thành phần hóa học giống collagen. HÌnh thành nền
tảng cấu trúc cho các cơ quan như lách và hạch bạch huyết.
- Các sợi collagen được tổ chức thành bó sợi
- Nguyên bào sợi được bao bọc bởi các sợi collagen
- Fibronectin là một protein ngoại bào đóng vai trò quan trọng hỗ trợ tế bào liên kết với chất nền ngoại
bào.
Ôn tập Tế bào học pg. 36
Nguyễn Minh Khôi
Có hai cách chủ yếu để các tế bào động vật tương tác với nhau
- Các sợi trung gian neo giữ tế bào với chất nền và tế bào với nhau tại vùng adhesion
- Chất nền ngoại bào chịu đựng trực tiếp các lực căng và nén
Biểu mô và liên kết tế bào
- Các tế bào có thể được “đóng gói” lại với nhau theo các cách khác nhau để hình thành các lớp biểu mô
*) Các loại liên kết tế bào trong cấu trúc mô động vật
- Liên kết neo giữ
+ Vùng gắn vi sợi: liên kết tế bào – tế bào (adherens do các bó vi sợi giữa hai tế bào tạo ra) và
liên kết tế bào – chất nền (actin – linked cell matrix adhesions)
+ Vùng gắn với sợi trung gian: liên kết tế bào – tế bào (desmosomes do các sợi trung gian chủ
yếu là keratin) và liên kết tế bào – chất nền (hemidesmosomes hay bán liên kết)
adheren và desmosome đều được xây dựng xung quanh các protein xuyên màng thuộc họ cadherin.
Cadherin đóng vai trò chính trong các liên kết phụ thuộc Ca2+ ở tế bào động vật.
Ôn tập Tế bào học pg. 37
Nguyễn Minh Khôi
Trong desmosomes, nhóm các cadherin kết nối với các sợi keratin (đặc trưng của biểu mô)
Các intergrin liên kết chất nền ngoại bào với bộ vi sợi và sợi trung gian
- Liên kết dải bịt
+ Liên kết chặt tight junction (trong động vật có xương sống) bịt các tế bào gần nhau trong lớp biểu
mô giúp lớp biểu mô chống rò rỉ và phân chia mặt trên và mặt đáy
+ Liên kết vách ngăn (trong động vật không xương sống)
- Liên kết tạo kênh
+ Gap junctions – liên kết khe (trong động vật) Cung cấp cho các tế bào gần nhau một kênh trực
tiếp (tạo bởi phức hệ protein xuyên màng 6 lần connexon ) để kết nối thông nhau, cho phép các phân tử tan
trong nước nhỏ trong hai tế bào trao đổi với nhau như các ion vô cơ và các sản phẩm chuyển hóa.
+ Plasmodesmata junction (trong thực vật)
- Liên kết chuyển tiếp tín hiệu
+ Các synap hóa học trong hệ thần kinh
+ Các synap miễn dịch trong hệ thống miễn dịch
+ Các thụ thể xuyên màng trong tiếp xúc tế bào – tế bào (Delta –Notch, ephrin – Eph…)
Ôn tập Tế bào học pg. 38
Nguyễn Minh Khôi
Liên kết neo giữ, dải bịt và tạo kênh cũng có thêm chức năng tín hiệu ngoài chức năng cấu trúc
- Các protein xuyên màng liên kết bộ khung tế bào với cấu trúc ngoại bào
Ba cơ chế tương tác trong liên kết tế bào
- Homophilic bingding: tương tác đặc hiệu như kháng nguyên – kháng thể
- Heterophilic binding: tương tác không đặc hiệu
- Liên kết thông qua một phân tử trung gian (kém phổ biến nhất)
CHU KỲ TẾ BÀO
- Chu kỳ tế bào là một vòng trong cuộc sống của nó bao gồm lớn lên, sao chép các nhiễm sắc thể, tách các
nhiễm sắc thế rồi phân chia…
- Sự phân chia tế bào là một chuỗi các sự kiện diễn ra trong một tế bào dẫn tới sự phân chia và sự nhân đôi
của nó…
- Thời gian của chu kỳ tế bào là rất khác nhau tùy loại tế bào: tế bào trong phôi sớm ếch – 30p; tế bào gan
người ~ 1 năm.
- Chu kỳ của tế bào nhân thực được chia là 4 pha:
+ Pha phân chia (mitosis)
+ Pha G1 (khoảng trống 1)
+ Pha S (tổng hợp) Kỳ trung gian hay Gian kỳ (Interphase)
+ Pha G2 (khoảng trống 2) chiếm 90% thời gian chu kỳ tế bào
- Chu kỳ tế bào được điều chỉnh bởi hệ thống phân tử điều khiển
+ Sự phân chia tế bào có diễn ra thường xuyên hay không còn phụ thuộc vào loại tế bào và tình
trạng của tế bào.
+ Những khác biệt về chu kỳ tế bào do sự điều chỉnh ở cấp độ phân tử
+ Chu kỳ tế bào được vận hành bởi các tín hiệu hóa học đặc trưng biểu hiện trong tế bào chất
Ôn tập Tế bào học pg. 39
Nguyễn Minh Khôi
+ Các cấp độ của các tín hiệu hóa học bị ảnh hưởng của các nhân tố sinh học hoặc phi sinh học.
Hệ thống điều khiển chu kỳ tế bào bấm nút cho các quá trình chính trong chu kỳ
- Hệ thống điều khiển các diễn biến trong cả chu kỳ thông qua 3 điểm chuyển tiếp
- Điều khiển chu kỳ tế bào là tương tự nhau ở các sinh vật nhân thực, về cơ bản sử dụng chung một cỗ máy
*) Hệ thống điều khiển chu kỳ tế bào phụ thuộc vào hoạt tính theo chu kỳ của protein kinase được gọi là
Cdks và nồng độ protein cyclin
- Hệ thống điều khiển chu kỳ tế bào hoạt động bằng cách hoạt hóa hoặc ức chế một cách có chu kỳ các
protein hay phức hệ protein then chốt thông qua quá trình phosphoryl (kinase) hay dephosphoryl
(phosphatase) hóa các protein này
- Các kinase có mặt trong suốt chu kỳ nhưng chỉ hoạt tính đúng lúc.
- Các kinase này có hoạt tính hay không phụ thuộc vào cyclin protein và vì thế các kinase này là cyclin –
dependent kinases (Cdks)
- Cyclin có nồng độ thay đổi theo chu kỳ tế bào, và mỗi khi nó có nồng độ cao đột biến, các Cdks được
hoạt hóa (cũng dựa vào quá trình phosphoryl hóa protein Cdks). Mặt khác, cyclins hướng các Cdk tới các
protein đích mà Cdk cần hoạt hóa.
Những phức hệ Cyclin – Cdk bấm nút cho các giai đoạn khác nhau trong chu kỳ tế bào
Ôn tập Tế bào học pg. 40
Nguyễn Minh Khôi
Nồng độ cyclin được điều chỉnh bởi sự phiên mã và sự thủy phân protein
- Trong suốt chu kỳ tế bào, nồng độ của mỗi cyclin tăng dần dần (có bản chất là dựa vào sự phiên mã các
gene cyclin) và sau đó giảm đột ngột (do sự phá hủy toàn bộ có tổ chức)
- Sự thoái biến của M và S cyclin phụ thuộc nhiều vào phức hệ enzym lớn là –anaphase – promoting
complex (APC).
- APC gắn các cyclin với một protein điều hòa là abuquitin. Khi đó cyclin ấy bị đưa tới proteasome và
thoái hóa nhanh chóng Cdks ngừng hoạt động.
Điều này lại nhắc ta một điều quan trọng trong tín hiệu tế bào: việc tắt các tín hiệu cũng quan trọng
tương tự như là việc bật nó lên.
Ôn tập Tế bào học pg. 41
Nguyễn Minh Khôi
Hoạt động của hệ cyclin – Cdk phụ thuộc vào sự phosphoryl hóa và dephosphoryl hóa
- Mặc dù nồng độ của cyclins tích lũy liên tục và tăng từ từ nhưng hoạt tính của Cdks lại tăng đột ngột vào
thời điểm thích hợp. Tại sao?
- Phức hệ Cyclin – Cdk chứa các phosphate Pi ức chế, và để trở nên có hoạt tính, Cdk phải được
dephosphoryl hóa bởi một protein phosphatase đặc hiệu.
- Hoạt động của Cdk có thể khóa bởi các protein ức chế Cdk (Cdk inhibitor proteins). Trong trường hợp
này, mặc dù cyclin – Cdk đã được hoạt hóa, tuy nhiên lại bị một protein ức chế bám vào
+ VD Cdk bị ức chế trong hầu hết G1 và trì hoãn vào S (để có thời gian tổng hợp đủ và chờ điều
kiện môi trường) do p27
Hệ thống điều khiển chu kỳ tế bào có thể tạm dừng chu kỳ bằng nhiều cách khác nhau
- Hệ thống có thể tạm dừng chu kỳ tại các checkpoint.
- Sử dụng kết hợp các cơ chế đã nêu.
Ôn tập Tế bào học pg. 42
Nguyễn Minh Khôi
*) Pha G1
- Là thời gian vội vã để trao đổi chất, phát triển tế bào, sửa chữa.
- Tiếp nhận các tín hiệu nội bào (về kích cỡ tế bào, hư hại DNA) và tín hiệu ngoại bào (về điều kiện môi
trường) để quyết định tới S hay dừng lại hoặc vào Go
Điểm chốt G1 -S là điểm chốt quan trọng nhất trong ba điểm chốt chu kỳ.
Cdks bị bất hoạt hoàn toàn trong G1
- Bất hoạt tất cả các M và S Cdks còn dư sau pha M bằng cách tìm phá các cyclin còn lại.
+ Ngăn chặn tổng hợp thêm cyclins mới
+ Phá hủy các cyclin còn lại
+ Triển khai các protein ức chế Cdks (theo cơ chế ở trên)
Mitogens đẩy mạnh sự sản xuất cyclins thúc đẩy phân chia tế bào
- Mitogens được sinh ra bởi một tế bào khác, thúc đẩy tế bào nào đó sinh sôi (ở đv có vú)
- Nếu tế bào không nhận được mitogens trong thời gian dài, nó sẽ chuyển sang Go
- Mitogens kích hoạt và thúc đẩy quá trình tổng hợp G1- cyclin, G1/S – cyclin và nhiều protein cần thiết
khác.
Sự hư hại DNA có thể dừng tạm thời tiến trình ở G1
- DNA hư hại ở G1 gây ra sự tăng cả về nồng độ và hoạt tính của một protein được gọi là p53
- p53 là một nhân tố điều hòa phiên mã cho gene mã hóa protein ức chế là p21
Ôn tập Tế bào học pg. 43
Nguyễn Minh Khôi
- Protein p21 bám vào phức hệ
G/S – Cdk và S/Cdk, ngăn cản
chúng đưa tế bào tới pha S
- Sự tạm dừng tại G1 cho tế
bào thời gian sửa DNA.
- Nếu DNA hỏng quá nặng,
p53 kích hoạt một lộ trình chết
tế bào apoptosis
- Sự thiếu hụt hay sai hỏng của
p53 đưa đến những khả năng
cao của đột biến và ung thư
Tế bào có thể trì hoãn việc phân chia trong một thời kỳ dài bằng cách đi vào một trang thái nghỉ đặc
biệt
- Tế bào có thể được đưa vào pha không phân chia (Go) một vài giờ, vài ngày…hay mãi mãi.
- Nhiều tế bào sẽ đi vào trạng thái ngừng phân chia Go vĩnh viễn sau lần biệt hóa cuối cùng của nó (tế bào
thần kinh, cơ). Khi đó, hệ thống điều khiển bị tháo dỡ vĩnh viễn, các gene mã hóa cyclin và Cdks bị tắt
hẳn không thể bật lại.
- Hầu như tính đa dạng về thời gian giữa hai lần phân chia ở các loại tế bào khác nhau là do G1 hay Go
PHA S – G2
S – Cdk bắt đầu quá trình sao chép và ngăn quá trình sao chép lặp lại nhiều lần
Sự sao chép không hoàn thành có thể giữ tế bào ở G2
- Tế bào cần đảm bảo rằng DNA đã sao chép hoàn toàn, đúng một lần và không có sai hỏng nào trước khi
sang M
- Như đã giới thiệu, M –Cdk bị ức chế do sự phosphoryl hóa các vị trí đặc hiệu. Và Cdc25 dephosphoryl
hóa nó để nó hoạt động lại khi phù hợp.
- Nếu các vấn đề xảy ra, tế bào bị giữ ở G2, Cdc25 tự ức chế nó cho tới khi vấn đề được giải quyết
PHA M
Ôn tập Tế bào học pg. 44
Nguyễn Minh Khôi
- Là thời gian ngắn nhất của 4 pha nhưng bùng nổ nhất
M – Cdk dẫn con đường đi vào pha M và Mitosis
- Phức hệ M – Cdk được tích lũy trong G2 nhưng đến tận cuối G2 mới được kích hoạt nhờ Cdc25
- M – Cdk được hoạt hóa có thể tạo ra hai phản hồi đến Cdc25 đang bị ức chế (kích hoạt lạiprotein này) và
các kinase ức chế Wee1 (tắt kinase này đi) để hoạt tính M – Cdk tăng mạnh, đưa tế bào tới M
-
Cohesins và Condensins giúp định hình các NST kép cho sự tách
- Tại pha M, các nhiễm sắc thể kép đặc lại, nhờ vào phức hệ protein condensin
- M – Cdk bấm nút kích hoạt (bằng phosphoryl hóa) cho condensin làm condensins bám vào nhiễm sắc thể
- Ngay cả khi đang còn trong pha S, các nhiễm sắc tử chị em được gắn chặt lấy nhau bởi phức hệ cohensin
dọc chiều dài của chuỗi. (Khuyết tật trong cohensin dẫn tới hội chứng Down)
Các dạng tập hợp của bộ khung tế bào thực hiện mitosis và cytokinesis
- Thoi vô sắc mitotic spindle là các vi ống tương tác với các protein đặc hiệu để tách các nhiễm sắc thể
kép
- Vòng thắt contractile ring gồm các sợi actin và các sợi trung gian có vài trò phân chia tế bào chất
Ôn tập Tế bào học pg. 45
Nguyễn Minh Khôi
Pha M xảy ra theo các bước
- Prophase
- Prometaphase Mitosis
- Metaphase
- Anaphase
- Telophase Cytokinesis
Ôn tập Tế bào học pg. 46
Nguyễn Minh Khôi
Các trung tử nhân lên để giúp hình thành hai cực của thoi phân bào
Ôn tập Tế bào học pg. 47
Nguyễn Minh Khôi
- Hai hiện tượng quan trọng trước khi pha M bắt đầu là DNA phải được nhân đôi hoàn toàn và trung tử
cũng nhân đôi. Hai quá trình đó bắt đầu cùng lúc, cùng bởi G1 – Cdk và G1/S – Cdk
- Khi được nhân đôi, hai trung tử nằm cùng chỗ.
Thoi phân bào bắt đầu hình thành ở Kỳ đầu (prophase)
- Các nhiễm sắc thể co xoắn và có thể quan sát dưới kính hiển vi
- 2 trung tử tách ra và tỏa ra các vi ống (tạo nên thể sao aster) trong khi đang di chuyển về 2 phía đối diện
- Thoi phân bào bắt đầu hình thành. Tính ổn định của các vi ống trên thoi phân bào ban đầu giảm do M –
Cdk thêm Pi vào các protein liên kết vi ống.
- Một số vi ống từ hai cực tương tác (nhờ các motor protein) để tăng tính bền vững gọi là interpolar
microtubules
Các nhiễm sắc thể gắn vào các thoi tơ vô sắc ở trước kỳ giữa (prometaphase)
- Tấm lamina bị phân giải, màng nhân tiêu biến, tan ra thành các bóng không bào nhỏ phân tán trong tế bào
chất. Hạch nhân nhỏ dần rồi biến mất
- Quá trình này được bấm nút bởi sự phosphoryl hóa các protein lỗ nhân và protein lamina.
- Nhân cô đặc hơn
- Các thoi tơ của thoi phân bào đang chờ sẵn tiếp cận các nhiễm sắc thể và bắt giữ chúng
- Thoi vô sắc bám vào NST tại các kinetochores. Mỗi chromatid có 1 kinetohore.
- Số lượng vi ống gắn vào mỗi kinetochore cò tùy thuộc loài, mỗi kinetochore của người bám với 20 – 40
vi ống, trong khi nấm men có mỗi 1.
- Đến thời điểm này, ta có rõ ràng ba loại vi ống khác nhau về chức năng: aster microtubules, kinetochore
microtubules và interpolar microtubules.
Các nhiễm sắc thể giúp hình thành thoi phân bào: trong các tế bào không có trung tử như tế bào động
vật hay một số tế bào động vật, các nhiễm sắc thể tập trung các vi ống, các motor protein sau đó di chuyển,
sắp xêp các vi ống và các nhiễm sắc thể thành một thoi lưỡng cực.
Chú ý rằng trong các tài liệu không có kỳ đầu giữa, nên các diễn biến của pha này sẽ thuộc về cuối kỳ
đầu.
Các nhiễm sắc thể xếp thành hàng trên xích đạo của thoi phân bào tại kỳ giữa (metaphase)
- Các nhiễm sắc thể xếp thành một hàng trên mặt phẳng xích đạo của thoi phân bào, hình thành metaphase
plate.
- Các vi ống vẫn tiếp tục kéo cả hai phía kinetochores và các nhiễm sắc thể đưa đẩy qua lại để điều chỉnh
vị trí của nó.
Sự thủy phân protein (protelysis) bấm nút cho quá trình tách nhiễm sắc tử chị em ở kỳ sau
(annaphase )
- Đột ngột cắt đứt các cohesin. Hai nhiễm sắc thể (giờ được coi như nhiễm sắc thể) có thể được kéo về hai
cực của thoi
- Cohesin bị cắt bởi protease là separase. Trước khi kỳ sau bắt đầu, enzym này bị ức chế bởi securin. Khi
bắt đầu kỳ sau,APC nhắm tới và phá hủy securin
Ôn tập Tế bào học pg. 48
Nguyễn Minh Khôi
Các nhiễm sắc thể vẫn tiếp tục tách ra trong suốt kỳ sau
- Tất cả các nhiễm sắc thể đồng thời đi về hai cực của tế bào với tốc độ đồng đều 1µm/min.
- Sự di chuyển là kết quả của anaphase A và anaphase B có thể cùng lúc hoặc không.
+ Trong anaphase A, kinetochore microtubules bị thu ngắn chủ yếu do sự giải trùng hợp ở cả hai
đầu
+ Trong anaphase B, hai cực của thoi tự tách xa nhau, lực sinh ra do các protein trong họ kinesin
và dynein. Kinesin hoạt động trên các interpolar microtubules di chuyển về chiều dương ; dynein bám vào
các aster microtubules và neo vào lớp vỏ tế bào kéo trung tử ra xa
Một nhiễm sắc thể không được gắn sẽ cản lại sự tách của các nhiễm sắc tử chị em
Ôn tập Tế bào học pg. 49
Nguyễn Minh Khôi
- Nếu việc gắn chưa hoàn thành, một lộ trình tín hiệu trong mitosis bật lên để chặn APC, các vòng cohesin
không tách ra
đây là spindle assembly checkpoint
Màng nhân được tái tạo ở kỳ cuối (telophase)
- Các nhiễm sắc thể di chuyển về hai cực của tế bào.
- Tháo dỡ thoi phân bào
- Tái lắp ghép màng nhân ở xung quanh mỗi nhóm nhiễm sắc thể con
- Các protein lỗ nhân và lamina nay được dephosphoryl hóa và tập hợp trở lại
- Khi có màng nhân, các lỗ nhân bơm các protein nhân làm phồng nhân
- Các nhiễm sắc thể giãn xoắn
kết thúc mitosis
Cytokinesis
- Thường bắt đầu từ kỳ sau và cho tới tận khi nhân của hai tế bào con hình thành ở kỳ cuối
Thoi phân bào xác đinh mặt phẳng phân cắt tế bào chất
- Mặt phẳng cắt luôn luôn vuông góc với trục của thoi phân bào, đảm bảo cắt chính giữa hai nhóm nhiễm
sắc thế đã phân chia.
- Tuy nhiên, không phải luôn luôn mặt phẳng này chia hai tế bào con có kích thước ngang nhau. Trong phát
triển phôi, thoi phân bào không nằm đối xứng, và tạo ra hai tế bào khác nhau về kích cỡ, cũng như các phân
tử mà chúng nhận được. Bởi chúng sẽ phát triển ra các loại tế bào khác nhau.
Contractile ring của các tế bào động vật được tạo nên từ các sợi actin và myosin
- Là một dải chồng gối của các sợi actin với các sợi myosin
- Chúng tập hợp tại kỳ sau và liên kết với các protein bám màng tại mặt trong của màng sinh chất
- Lực được sinh ra khi các actin trượt qua myosin
- Contractile là một cấu trúc tạm thời, vòng này nhỏ dần và biến mất khi hai tế bào đã tách nhau ra
Cytokinesis ở tế bào thực vật đòi hỏi sự hình thành thành tế bào
- Chúng không có contratile ring
- Thành tế bào bắt đầu hình thành ở giữa hai bộ nhiễm sắc thể đang phân li ở đầu kỳ cuối, đó là cấu trúc
phragmoplast tạo thành do các interpolar microtubules còn lại.
- Các túi polisaccaride và glycoprotein từ Golgi được đưa đến phragmoplast (theo các vi ống)
- Chúng hình thành một cấu trúc dạng đĩa, các bóng không bào tiếp tục tan ra để đĩa mở rộng cho tới khi
nó chạm tới màng tế bào có sẵn
- Cuối cùng các sợi cellulos được đặt vào thành thành tế bào.
Các bào quan có màng phải được phân bố cho các tế bào con trong phân chia tế bào
ĐIỀU KHIỂN SỐ LƯỢNG VÀ KÍCH THƯỚC TẾ BÀO
Apoptosis giúp điều chỉnh số lượng tế bào động vật
- Apoptosis là một chương trình chết tế bào
- Hết các tế bào động vật tự tử nhờ lộ trình apoptosis
- Số lượng tế bào cần được kiểm soát nghiêm ngặt và hệ thống điều khiển chy kỳ tế bào thôi là chưa đủ.
Sinh phải đi liền với tử. Sự chết tế bào thường cân đối một cách chính xác với sự phân chia tế bào.
Apoptosis được trung gian bởi một lớp tác nhân thủy phân protein (Proteolytic) nội bào
- Khác với cái chết hoại tử (necrosis)do tai nạn, chết bằng apoptosis rất gọn gàng
- Ban đầu tế bào xuất hiện các bọng nước (blebs) không đều trên bề mặt, sau đó co lại và đặc hơn. Bộ khung
xương tế bào sụp đổ, màng nhân tan biến, DNA nhân vỡ thành nhiều mảnh vụn.
Ôn tập Tế bào học pg. 50
Nguyễn Minh Khôi
- Các tế bào này lập tức thu hút các đại thực bào ; đại thực bào lập tức bao bọc tất cả thể apoptotic này (giờ
nó không còn là một tế bào nữa rồi) trước khi đánh sập nó
-
- Một họ protease có liên quan chặt chẽ tới apoptosis là caspases, khi chưa được kích hoạt chúng gọi là
procaspase
- Có hai loại caspase hoạt động cùng nhau : initiator caspases( caspases mở đầu) thực hiện cắt và từ đó
hoạt hóa, kéo theo excutioner caspases (caspases hành hình). Chúng đều có khả năng phản hồi lại yếu tố
hoạt hóa để khuếch đại tín hiệu. Các caspases này thuộc họ protein thủy phân
Bên trong Apoptotic được điều khiển bởi họ protein nội bào Bcl2
- Bên trong nhân của tất cả các tế bào động vật chứa đựng những hạt giống của chính sự phá hủy :
procaspases.
- Một họ protein nội bào giám sát hoạt tính caspases là Bcl2, họ này bao gồm các thành viên thúc đẩy hoạt
tính caspase và sự chết tế bào, trong khi các thành viên khác ức chế điều đó. Tiêu biểu là Bac và Bax
+ Các protein này hoạt hóa để đáp ứng tín hiệu phát do khi có hư hại DNA và các vấn đề khác,
chúng thúc đẩy quá trình giải phóng chất dẫn truyền điện tử cytochrome c từ ty thể ra tế bào chất
+ Các thành viên khác của họ Bcl2 (bao gồm một phân tử cũng có tên Bcl2) ức chế apoptosis bằng
cách chặn Bax và Bac giải phóng cytochrom c
- Các phân tử cytochrom c được giải phóng từ ty thể hoạt hóa các procaspases, sau đó thúc đẩy sự tập hợp
phức hệ protein lớn 7 tay được gọi là apoptosome. Apoptosome tìm bắt các procaspase đặc trưng, để kích
hoạt một lớp các caspases dẫn tới apoptosis
Tín hiệu ngoại bào cũng có thể dẫn tới apoptosis
- Có một bộ các protein thụ thể bền mặt màng tế bào được coi là death receptors. Tiêu biểu phải kể đến
Fas. Lộ trình này như sau :
+ Fas ligand là một protein bám màng trên các bạch cầu sát thủ (killer lymphocyte)
+ Fas ligand bám vào Fas trên màng tế bào cần phải chết, làm kích hoạt lộ trình apoptosis bên trong tế bào
đó
Ôn tập Tế bào học pg. 51
Nguyễn Minh Khôi
Các tế bào đòi hỏi các tín hiệu ngoại bào để tồn tại, sinh trưởng và phân chia
- Survival factors thúc đẩy tế bào tồn tại, rộng hơn là ngăn ngừa apoptosis
- Mitogens thúc đẩy tế bào phân chia, chủ yếu bằng cách vượt qua cơ chế phanh nội bào có xu hướng ngăn
các quá trình trong chu kỳ tế bào.
- Growth factors thúc đẩy sự phát triển tế bào (Tăng kích thươc tế bào và sinh khối) bằng cách khuyến
khích chuyển hóa và ngăn chặn sự thoái biến của các protein và các đại phân tử khác
Các nhân tố tồn tại ngăn chặn apoptoisis
- Các growth factors thường hoạt hóa các thụ thể bề mặt tế bào để ức chế họ Bcl2, chặn lộ trình apoptosis
Mitogens đẩy mạnh sự phân chia tế bào bằng cách thúc đẩy đi vào pha S
- Lưu ý tới nhân tố tiêu biểu là nhân tố sinh trưởng từ tiểu cầu (platelet-derived growth factor, or
PDGF)
Các nhân tố sinh trưởng thúc đẩy phân chia tế bào
- Dẫn tới tích lũy các đại phân tử và protein. Đẩy nhanh quá trình tổng hợp và kìm chế sự thoái biến các
phân tử này
Một số protein tín hiệu ngoại bào ức chế sự sống sót, phân chia và phát triển
Ôn tập Tế bào học pg. 52
You might also like
- Uống nước sinh tố: Phương pháp kỳ diệu - bảo vệ sức khỏe và trị liệu bệnh tật.From EverandUống nước sinh tố: Phương pháp kỳ diệu - bảo vệ sức khỏe và trị liệu bệnh tật.No ratings yet
- Đề cương tế bàoDocument43 pagesĐề cương tế bàonot? WhyNo ratings yet
- Đề Cương Sinh Học Kì IDocument8 pagesĐề Cương Sinh Học Kì IAnh TuanNo ratings yet
- Sinh Lý Các Bào QuanDocument4 pagesSinh Lý Các Bào QuanNghiem NguyenNo ratings yet
- SinhDocument7 pagesSinhHoàng Vinh NguyễnNo ratings yet
- SINHHK1Document5 pagesSINHHK1Quân đoànNo ratings yet
- ĐỀ CƯƠNG SINH CUỐI KÌ 1Document7 pagesĐỀ CƯƠNG SINH CUỐI KÌ 1Minh NguyễnNo ratings yet
- Tế bào nhân thưucjDocument3 pagesTế bào nhân thưucjVũ Tâm MinhNo ratings yet
- ÔN TẬP SHTB Tóm TắtDocument17 pagesÔN TẬP SHTB Tóm TắtTrương Gia Bảo0% (1)
- Sinh học PTTB Nhân sơ nhân thực, vận chuyển bằng túiDocument18 pagesSinh học PTTB Nhân sơ nhân thực, vận chuyển bằng túinguyentin.cao21104No ratings yet
- Te Bao Eukaryote - 3Document15 pagesTe Bao Eukaryote - 3Thục Bình NguyễnNo ratings yet
- CHUYÊN ĐỀ 3Document29 pagesCHUYÊN ĐỀ 3Quang Long LươngNo ratings yet
- 02.2 - EukaryoteDocument99 pages02.2 - EukaryoteLý KỳNo ratings yet
- SinhDocument7 pagesSinhHai ĐăngNo ratings yet
- Tự Luận SinhDocument5 pagesTự Luận SinhHoàng HảiNo ratings yet
- đề cương sinh học mớiDocument7 pagesđề cương sinh học mớiVũ Mỹ HuyềnNo ratings yet
- Bài 3. Tê Bào - SV - 2022Document52 pagesBài 3. Tê Bào - SV - 2022Cuong NguyenNo ratings yet
- Đề Cương Sinh HọcDocument3 pagesĐề Cương Sinh HọcHarley Quinn MelanieNo ratings yet
- Tế Bào Nhân ThựcDocument39 pagesTế Bào Nhân ThựcThịnh ĐinhNo ratings yet
- Sinh Đ I Cương 2Document184 pagesSinh Đ I Cương 203.Trần Mai Trâm AnhNo ratings yet
- Chương 1-5 H NGDocument45 pagesChương 1-5 H NGMai HồngNo ratings yet
- ĐỀ CƯƠNG SINH 10 HSDocument3 pagesĐỀ CƯƠNG SINH 10 HSDanh ThânNo ratings yet
- Ôn Lý Thuyết (Rất Đủ)Document32 pagesÔn Lý Thuyết (Rất Đủ)gg2minutesNo ratings yet
- Tế Bào Nhân Sơ, Nhân Thực (Pkn)Document12 pagesTế Bào Nhân Sơ, Nhân Thực (Pkn)Ziee NqọcNo ratings yet
- Đáp Án 10 Nâng CaoDocument25 pagesĐáp Án 10 Nâng CaoZaP NguyenNo ratings yet
- Noi Dung Mô PhôiDocument47 pagesNoi Dung Mô PhôiNguyễn Nam KhánhNo ratings yet
- Cau Truc Te BaoDocument23 pagesCau Truc Te BaoNam LeNo ratings yet
- Sinh học lớp 10 Bài 8Document9 pagesSinh học lớp 10 Bài 8Hạnh Nguyên Trần NguyễnNo ratings yet
- slide sinh học tế bàoDocument40 pagesslide sinh học tế bàoNguyễn Dương HuyNo ratings yet
- Bai 8 Te Bao Nhan ThucDocument15 pagesBai 8 Te Bao Nhan ThucHoàng Kim Anh PhụngNo ratings yet
- Copy of MÀNG SINH CHẤT - VẬN CHUYỂN CÁC CHẤT QUA MÀNGDocument23 pagesCopy of MÀNG SINH CHẤT - VẬN CHUYỂN CÁC CHẤT QUA MÀNGLuong HaNo ratings yet
- Đáp án chỉnh sửa - Sinh HọcDocument16 pagesĐáp án chỉnh sửa - Sinh HọcTran SangNo ratings yet
- CÂU HỎI ÔN TẬP TỰ LUẬN HỌC KỲ 1Document2 pagesCÂU HỎI ÔN TẬP TỰ LUẬN HỌC KỲ 1toplqvn2k8No ratings yet
- Cau Truc Te Bao Nhan Thuc - Minh Tien 10 Hoa LNDocument2 pagesCau Truc Te Bao Nhan Thuc - Minh Tien 10 Hoa LNMinh Tiến NguyễnNo ratings yet
- Đề Cương SinhDocument16 pagesĐề Cương SinhPhan YunaNo ratings yet
- Đề Cương Sinh Học VVHDocument28 pagesĐề Cương Sinh Học VVHNhi LêNo ratings yet
- 1. Vận chuyển vật chất qua màng tế bàoDocument19 pages1. Vận chuyển vật chất qua màng tế bàoThanh HằngNo ratings yet
- Sinh họcDocument78 pagesSinh họcNVTNo ratings yet
- Đề cương sinh 2Document8 pagesĐề cương sinh 2Phạm Thu NgàNo ratings yet
- Bai Giang TE BAO PDFDocument9 pagesBai Giang TE BAO PDFTrần Duy AnhNo ratings yet
- Lưới nội chất - Bộ máy GolgiDocument12 pagesLưới nội chất - Bộ máy Golginguyễn gia hưng phạmNo ratings yet
- Sinh 10 hk1Document20 pagesSinh 10 hk1Vũ HoàngNo ratings yet
- Hinh Thai Giai Phau Thuc VatDocument59 pagesHinh Thai Giai Phau Thuc VatMatalus MasterNo ratings yet
- B7,10 Sinh Cánh diềuDocument3 pagesB7,10 Sinh Cánh diềuKim NgọcNo ratings yet
- BT SinhDocument8 pagesBT SinhVydzNo ratings yet
- SINH- - - - - - I-C - - - - NG-V - - -DI-TRUY - - - N-ch - - - - ng-1.pptx; filename= UTF-8''SINH-ĐẠI-CƯƠNG-VÀ-DI-TRUYỀN-chương-1Document117 pagesSINH- - - - - - I-C - - - - NG-V - - -DI-TRUY - - - N-ch - - - - ng-1.pptx; filename= UTF-8''SINH-ĐẠI-CƯƠNG-VÀ-DI-TRUYỀN-chương-1Desu HeiNo ratings yet
- Sinh Hoc Te BaoDocument24 pagesSinh Hoc Te Baomainguyentranxuan11b221No ratings yet
- Bài 9 - Tế bào nhân thựcDocument7 pagesBài 9 - Tế bào nhân thựcÁi ThơNo ratings yet
- Phan 2 Sinhhocdaicuong 2023Document184 pagesPhan 2 Sinhhocdaicuong 2023sky14112005No ratings yet
- Chương 2Document3 pagesChương 2linhdao1901vnNo ratings yet
- Sinh Học Tb-di (Đề Cương Kthp)Document107 pagesSinh Học Tb-di (Đề Cương Kthp)tung.lt.23yk1No ratings yet
- Kiến thức chương 5 - sinh 10Document4 pagesKiến thức chương 5 - sinh 10le4315514No ratings yet
- Bai 1Document45 pagesBai 1quill1206No ratings yet
- Đáp án hướng dẫn ôn tập sinh học đại cươngDocument25 pagesĐáp án hướng dẫn ôn tập sinh học đại cươngNT Thanh TuyềnNo ratings yet
- 3.1. de 1Document5 pages3.1. de 1Roam ResearcherNo ratings yet
- BÀI 2. Cấu Trúc Và Các Hoạt Động Của TBDocument138 pagesBÀI 2. Cấu Trúc Và Các Hoạt Động Của TBPhương Quỳnh HoàngNo ratings yet
- Ôn tập cơ sở KHTNDocument11 pagesÔn tập cơ sở KHTNBảo TrâmNo ratings yet
- TẾ BÀO NHÂN THỰCDocument2 pagesTẾ BÀO NHÂN THỰCLe Quan Binh B2107563No ratings yet
- Chương 1: Biểu môDocument20 pagesChương 1: Biểu môVi Triệu TườngNo ratings yet
- ĐỀ CƯƠNG CUỐI KÌ 1 MÔN SINH 10Document26 pagesĐỀ CƯƠNG CUỐI KÌ 1 MÔN SINH 10mewmew4790No ratings yet