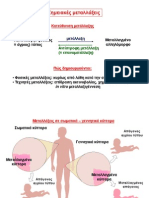
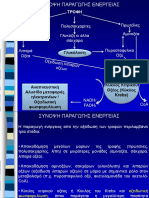
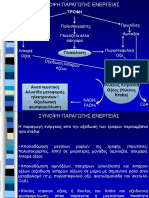
You might also like
- Ιατρική Μικροβιολογία Ι: Παθογόνα και ανθρώπινα μικροβιοκτόναFrom EverandΙατρική Μικροβιολογία Ι: Παθογόνα και ανθρώπινα μικροβιοκτόναNo ratings yet
- Βασική Γενετική Αμαλία-Σοφία Αφένδρα Πανεπιστήμιο ΙωαννίνωνDocument316 pagesΒασική Γενετική Αμαλία-Σοφία Αφένδρα Πανεπιστήμιο ΙωαννίνωνpapatsakalosNo ratings yet
- Η Βιολογία Κατεύθυνσης Γ' Λυκείου Με Αριθμούς-Φράσκου ΠαναγιώταDocument4 pagesΗ Βιολογία Κατεύθυνσης Γ' Λυκείου Με Αριθμούς-Φράσκου ΠαναγιώταPesmou TonponosouNo ratings yet
- 1 Σημειώσεις Οργάνωση Της Ζωής Β Γ ΓυμνασίουDocument2 pages1 Σημειώσεις Οργάνωση Της Ζωής Β Γ ΓυμνασίουSophia CooperNo ratings yet
- ΚΕΦ 1 ΠΙΝΑΚΕΣ ΔΙΑΦΟΡΩΝDocument16 pagesΚΕΦ 1 ΠΙΝΑΚΕΣ ΔΙΑΦΟΡΩΝStella SalvatoreNo ratings yet
- Απαντήσεις βιβλίου επαν κεφ1Document10 pagesΑπαντήσεις βιβλίου επαν κεφ1kyfkyfNo ratings yet
- Βιολογία κατεύθυνσης - Γ΄ Λυκείου - Απαντήσεις - ΟΕΦΕ 2003Document9 pagesΒιολογία κατεύθυνσης - Γ΄ Λυκείου - Απαντήσεις - ΟΕΦΕ 2003kljljklajjkjdljjflkjNo ratings yet
- ΒΙΟΛΟΓΙΑ Γ ΘΕΤΙΚΗ ΚΕΦ. 1Document6 pagesΒΙΟΛΟΓΙΑ Γ ΘΕΤΙΚΗ ΚΕΦ. 1bill80% (5)
- ΘΕΩΡΙΑ TEXN.AN.DNADocument7 pagesΘΕΩΡΙΑ TEXN.AN.DNAΜαρια ΞενουNo ratings yet
- Akadamia Proteinomena Themata 2014 Viologia Katefthinsis G LykeiouDocument12 pagesAkadamia Proteinomena Themata 2014 Viologia Katefthinsis G LykeiouAIMILIANo ratings yet
- ΒΙΟΛΟΓΙΑ ΝΑΣΙΑDocument257 pagesΒΙΟΛΟΓΙΑ ΝΑΣΙΑΓιάννης ΠαπαδόπουλοςNo ratings yet
- Πρόγραμμα Χαρτογράφησης Του Γονιδιώματος Του ΑνθρώπουDocument39 pagesΠρόγραμμα Χαρτογράφησης Του Γονιδιώματος Του ΑνθρώπουPinelopi PouliNo ratings yet
- σημειακές μεταλλάξειςDocument56 pagesσημειακές μεταλλάξειςΜαρία-Άννα ΚιρμπάκηNo ratings yet
- κυτταρα σωματος PDFDocument39 pagesκυτταρα σωματος PDFNikos PapamichailNo ratings yet
- Erwtiseis Krisews 2ou Kefalaiou KatefhinsisDocument8 pagesErwtiseis Krisews 2ou Kefalaiou KatefhinsisDery MourtziNo ratings yet
- New ΚΕΦΑΛΑΙΟ 4 -ΓΕΝΕΤΙΚΗDocument50 pagesNew ΚΕΦΑΛΑΙΟ 4 -ΓΕΝΕΤΙΚΗAthina MarkouNo ratings yet
- 1-Εργ.Βιολογίας Κυτταρο οργάνωση δομη-newDocument58 pages1-Εργ.Βιολογίας Κυτταρο οργάνωση δομη-newJimNo ratings yet
- Dialeksi 3Document69 pagesDialeksi 3giannisNo ratings yet
- Εισαγωγή Στη Βιολογία Ιωάννης Παυλίδης Πανεπιστημιο ΚρητηςDocument365 pagesΕισαγωγή Στη Βιολογία Ιωάννης Παυλίδης Πανεπιστημιο ΚρητηςpapatsakalosNo ratings yet
- Βιολογία κατεύθυνσης - Γ΄ Λυκείου - θέματα - ΟΕΦΕ 2003Document3 pagesΒιολογία κατεύθυνσης - Γ΄ Λυκείου - θέματα - ΟΕΦΕ 2003kljljklajjkjdljjflkj0% (1)
- 4. ΑναπαραγωγήDocument66 pages4. ΑναπαραγωγήathmastorakiNo ratings yet
- ΔΙΑΓΩΝΙΣΜΑ ΒΙΟΛΟΓΙΑΣ Β ΛΥΚΕΙΟΥ ΙΑΝΟΥΑΡΙΟΣ 2017 ΚΕΦ 1 2Document3 pagesΔΙΑΓΩΝΙΣΜΑ ΒΙΟΛΟΓΙΑΣ Β ΛΥΚΕΙΟΥ ΙΑΝΟΥΑΡΙΟΣ 2017 ΚΕΦ 1 2Anna SpanouNo ratings yet
- Το εσωτερικό του κυττάρουDocument19 pagesΤο εσωτερικό του κυττάρουapi-3730925100% (1)
- 4.2 Μοριακή ΓενετικήDocument18 pages4.2 Μοριακή ΓενετικήkwlosonNo ratings yet
- ΒΙΟΛΟΓΙΑ Γ΄ ΓΥΜΝΑΣΙΟΥDocument26 pagesΒΙΟΛΟΓΙΑ Γ΄ ΓΥΜΝΑΣΙΟΥMenelaos PanagiotouNo ratings yet
- Ce98 Ce9d 1cebf Ce91ce9dce91 Ce98ce95ce9cce91Document25 pagesCe98 Ce9d 1cebf Ce91ce9dce91 Ce98ce95ce9cce91dropbox623No ratings yet
- Γενετική Μαυραγάνη-Τσιπίδου Πηνελόπη ΑΠΘDocument124 pagesΓενετική Μαυραγάνη-Τσιπίδου Πηνελόπη ΑΠΘpapatsakalosNo ratings yet
- ΜΟΡΙΑΚΗ ΒΙΟΛΟΓΙΑ ΣΤΗ Μ-ΓDocument59 pagesΜΟΡΙΑΚΗ ΒΙΟΛΟΓΙΑ ΣΤΗ Μ-ΓMaradonaNo ratings yet
- Diag. Bio GlykeiouDocument6 pagesDiag. Bio Glykeiouneo212121No ratings yet
- ΜΑΘΗΜΑ 3Document57 pagesΜΑΘΗΜΑ 3Dr VenNo ratings yet
- ΓΙΑΒΑΣΗΣ ΓΕΝΙΚΗ ΜΙΚΡΟΒΙΟΛΟΓΙΑ 2020Document163 pagesΓΙΑΒΑΣΗΣ ΓΕΝΙΚΗ ΜΙΚΡΟΒΙΟΛΟΓΙΑ 2020JimNo ratings yet
- Βιολογία Κατεύθυνσης-Γ΄Λυκείου-Απαντήσεις στις ερωτήσεις του σχολικούDocument39 pagesΒιολογία Κατεύθυνσης-Γ΄Λυκείου-Απαντήσεις στις ερωτήσεις του σχολικούDemosthenes Karyophyllis69% (42)
- 1.3 Οργάνωση του γενετικού υλικού BWDocument21 pages1.3 Οργάνωση του γενετικού υλικού BWGeorgeSavvidisNo ratings yet
- DNA BarcodingDocument17 pagesDNA BarcodingPinelopi PouliNo ratings yet
- ΒΙΟΛΟΓΙΑ Α ΓΥΜΝΑΣΙΟΥDocument20 pagesΒΙΟΛΟΓΙΑ Α ΓΥΜΝΑΣΙΟΥOrestis Lazaridis100% (1)
- ΚΕΦ 1 ΟΡΟΛΟΓΙΑDocument7 pagesΚΕΦ 1 ΟΡΟΛΟΓΙΑStella SalvatoreNo ratings yet
- Βιολογία Κατεύθυνσης-Μάιος-2010Document14 pagesΒιολογία Κατεύθυνσης-Μάιος-2010Demosthenes KaryophyllisNo ratings yet
- Κεφ. 1 ΓΕΝΙΚΗ ΕΠΙΣΚΟΠΗΣΗ Α-Β PDFDocument40 pagesΚεφ. 1 ΓΕΝΙΚΗ ΕΠΙΣΚΟΠΗΣΗ Α-Β PDFNick LexisNo ratings yet
- Αρχές ανοσολογίαςDocument51 pagesΑρχές ανοσολογίαςsprfyllisNo ratings yet
- Ιοί Ιοειδή Prions (Ιατρική Οδοντιατρική)Document40 pagesΙοί Ιοειδή Prions (Ιατρική Οδοντιατρική)Emmanouil YamaNo ratings yet
- Antiviotiki Drasi AntoxiDocument76 pagesAntiviotiki Drasi Antoxiangela perNo ratings yet
- Bio 220112 PDFDocument2 pagesBio 220112 PDFKostas PachnisNo ratings yet
- ΕΡ. ΟΔΗΓΟΣ ΓΕΝΕΤΙΚΗΣ 2021Document53 pagesΕΡ. ΟΔΗΓΟΣ ΓΕΝΕΤΙΚΗΣ 2021Ευαγγελία ΓNo ratings yet
- ΒΙΟΛΟΓΙΑ Γ ΘΕΤΙΚΗ, ΚΕΦ. 1 - ΟρολογίαDocument2 pagesΒΙΟΛΟΓΙΑ Γ ΘΕΤΙΚΗ, ΚΕΦ. 1 - Ορολογίαbill100% (1)
- Εισαγωγή στη Μοριακή Βιολογία PDFDocument303 pagesΕισαγωγή στη Μοριακή Βιολογία PDFDimitris Katsidoniotis100% (1)
- 4.themata Exetasewn Anasyndyasmeno DNA CompressedDocument28 pages4.themata Exetasewn Anasyndyasmeno DNA Compressedmaria fermaniNo ratings yet
- 4.themata Exetasewn Anasyndyasmeno DNA CompressedDocument28 pages4.themata Exetasewn Anasyndyasmeno DNA CompressedΜαρία ΦερμάνηNo ratings yet
- ΒΙΟΛΟΓΙΑ Γ ΓΥΜΝ 5Document6 pagesΒΙΟΛΟΓΙΑ Γ ΓΥΜΝ 5Giannis LentzosNo ratings yet
- 395214374 ΒΙΟΛΟΓΙΑ Γ ΓΥΜΝ 5Document6 pages395214374 ΒΙΟΛΟΓΙΑ Γ ΓΥΜΝ 5bloubloubabalouNo ratings yet
- Κύτταρα Και Γονιδιώματα ΒιολογιαDocument12 pagesΚύτταρα Και Γονιδιώματα Βιολογιαarthra2No ratings yet
- Θέματα Πανελλαδικών 2000-2013, ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΑΝΑ ΚΕΦΑΛΑΙΟDocument98 pagesΘέματα Πανελλαδικών 2000-2013, ΒΙΟΛΟΓΙΑ ΘΕΤΙΚΗΣ ΑΝΑ ΚΕΦΑΛΑΙΟbiologismoi100% (1)
- 49 11 1o MathimaDocument89 pages49 11 1o MathimabiomedlabsciNo ratings yet
- 01. Εισαγωγή στη Μικροβιολογία ΤροφίμωνDocument74 pages01. Εισαγωγή στη Μικροβιολογία ΤροφίμωνTsosmiNo ratings yet
- Απαντημένες Ερωτήσεις ΓενετικήςDocument34 pagesΑπαντημένες Ερωτήσεις Γενετικήςanon_5707294No ratings yet
- Γενική Μικροβιολογία Και Μικροβιολογία Τροφίμων Ιωάννης Γιαβάσης Πανεπιστήμιο Θεσσαλίας 2019Document167 pagesΓενική Μικροβιολογία Και Μικροβιολογία Τροφίμων Ιωάννης Γιαβάσης Πανεπιστήμιο Θεσσαλίας 2019papatsakalosNo ratings yet
- 6) Ανοσία και φλεγμονήDocument90 pages6) Ανοσία και φλεγμονήPakiza ZhomartNo ratings yet
- Βιολογία Γ΄Λυκείου (Γ.Π.) Ειδική άμυναDocument32 pagesΒιολογία Γ΄Λυκείου (Γ.Π.) Ειδική άμυναDemosthenes Karyophyllis100% (1)
- 1. Μεταλλάξεις & Πολυμορφισμοί PDFDocument29 pages1. Μεταλλάξεις & Πολυμορφισμοί PDFΝενα ΧρονοπουλουNo ratings yet
- A-Risk Assessment 2017 NotesDocument6 pagesA-Risk Assessment 2017 NotesMary-jane VlsmNo ratings yet
- Air Quality and HealthDocument31 pagesAir Quality and HealthMary-jane VlsmNo ratings yet
- ΙΑΤΡΙΚΗ ΦΥΣΙΚΗ - ΦΥΣΙΚΗ ΤΟΥ ΑΝΘΡΩΠΙΝΟΥ ΣΩΜΑΤΟΣ - ΣΗΜΕΙΩΣΕΙΣ - 1 corrDocument134 pagesΙΑΤΡΙΚΗ ΦΥΣΙΚΗ - ΦΥΣΙΚΗ ΤΟΥ ΑΝΘΡΩΠΙΝΟΥ ΣΩΜΑΤΟΣ - ΣΗΜΕΙΩΣΕΙΣ - 1 corrMary-jane Vlsm100% (2)
- ΑΥΤΟΑΝΟΣΕΣ ΜΥΟΠΑΘΕΙΕΣ Γ. ΠΑΠΑΔΗΜΑΣDocument46 pagesΑΥΤΟΑΝΟΣΕΣ ΜΥΟΠΑΘΕΙΕΣ Γ. ΠΑΠΑΔΗΜΑΣMary-jane VlsmNo ratings yet
- Biostat - Environment&Health - 16-17Document132 pagesBiostat - Environment&Health - 16-17Mary-jane VlsmNo ratings yet
- Κεφάλαιο 7 ΑκτινοσκόπησηDocument23 pagesΚεφάλαιο 7 ΑκτινοσκόπησηNektaria LouiseNo ratings yet
- ΜαστογραφίαDocument16 pagesΜαστογραφίαMary-jane VlsmNo ratings yet
- ΔΙΑΛΕΞΕΙΣ 27η-29ηDocument348 pagesΔΙΑΛΕΞΕΙΣ 27η-29ηMary-jane VlsmNo ratings yet
- ΔΙΑΛΕΞΕΙΣ 23η-24ηDocument216 pagesΔΙΑΛΕΞΕΙΣ 23η-24ηMary-jane VlsmNo ratings yet
- ΔΙΑΛΕΞΕΙΣ 20η-22η σημDocument27 pagesΔΙΑΛΕΞΕΙΣ 20η-22η σημMary-jane VlsmNo ratings yet
- ΔΙΑΛΕΞΕΙΣ 25η-26ηDocument261 pagesΔΙΑΛΕΞΕΙΣ 25η-26ηMary-jane VlsmNo ratings yet
- ΔΙΑΛΕΞΕΙΣ 1η-3ηDocument321 pagesΔΙΑΛΕΞΕΙΣ 1η-3ηMary-jane VlsmNo ratings yet
- ΔΙΑΛΕΞΕΙΣ 15η-16η 2020Document53 pagesΔΙΑΛΕΞΕΙΣ 15η-16η 2020Mary-jane VlsmNo ratings yet
- Μεταβολισμός λιπιδίων PDFDocument24 pagesΜεταβολισμός λιπιδίων PDFMary-jane VlsmNo ratings yet
- Ευκαρυωτικό 2019Document48 pagesΕυκαρυωτικό 2019Mary-jane VlsmNo ratings yet
- Αναπνευστική αλυσίδα PDFDocument26 pagesΑναπνευστική αλυσίδα PDFMary-jane VlsmNo ratings yet
- Μεταβολισμός λιπιδίων PDFDocument24 pagesΜεταβολισμός λιπιδίων PDFMary-jane VlsmNo ratings yet
- Αναπνευστική αλυσίδα PDFDocument26 pagesΑναπνευστική αλυσίδα PDFMary-jane VlsmNo ratings yet
- MyobravoDocument47 pagesMyobravoMary-jane VlsmNo ratings yet