Professional Documents
Culture Documents
יחידה8
Uploaded by
Zohar ShaharabaniCopyright
Available Formats
Share this document
Did you find this document useful?
Is this content inappropriate?
Report this DocumentCopyright:
Available Formats
יחידה8
Uploaded by
Zohar ShaharabaniCopyright:
Available Formats
יחידה - 8נוקלאוטידים בס"ד
פנטוז )( 2
יחידה – 8מבנה ומטבוליזם של
פוספסט )( 3 נוקלאוטידים
הבסיסים החנקניים הם תוצרים של שני
קבוצות יסוד :פירמידינים ופורינים .שיטת מבוא והקדמה .1
המספור מתבססת על מספור הפחמנים והחנקן שבבסיס ,כדי להגדיר את לנוקלאוטידים תפקידים רבים במטבוליזם של התא :הם אחראים לתמורות
התוצרים ,וכן נקבע שם הנוקלאוטיד על פי הבסיס החנקני .כידוע ,הן ה האנרגיה בתא ,לקשרים הכימים הדרושים לתגובה נכונה של התא
DNAוהן ה RNAמכילים שני בסיסים פוריניים :אדנין ) (Aוגואנין ),(G להורמונים וגירויים מבחוץ ולכל מערך הרכיבים הדרושים לפעילותם
ובסיסים פירמדיניים :ציטוזין ) ,(Cוטימין ) (Tב DNAואורציל ) (Uב .RNA התקינה של אנזימים בתא .כמובן ,מעל לכל הם הבסיס לחומצות הגרעין:
לטבעות מבנה מוכר של קשרים כפולים ,ומשמעות הדבר שיש למולקולת חומצה דאואוקסיריבונוקלית ) (DNAוחומצה ריבונוקלית ) ,(RNAשהם
רזוננס ,וממילא מבנה כמעט פלנרי ,וכן בליעה חזקה של .UV היסודות המולוקולריים של המטען הגנטי .המבנה של כל חלבון ומרכיב
מבחינת מולקולות פנטוז ,ה DNAכולל -2דאוקסי-D-ריבוז וה RNAכולל המיוצרים בתא מקודד במטען התורשתי בעזרת חומצות הגרעין.
-Dריבוז; שתיהן מולקולת סוכר במצב מעגלי )בתרשים אנו רואים ריבוז ,כי המקטע ב DNAשמספק את המידע הדרוש לסנתוז של חלבון או RNA
יש לו קבוצת ] OHבאדום[ ,אם במקום קבוצת ה OHהיה רק ,Hזה נקרא גן .באופן טבעי מולקולות DNAהן גדולות מאוד משום שכל תא מכיל
הדאוקסי-ריבוז(. אלפים רבים של גנים .אחסון והעברת מידע גנטי הוא השימוש היחיד הידוע
מבנה הנוקלאוטיד :הבסיס של הנוקלאוטיד )הפירמידין או הפורין( מחובר לנו של ה .DNA
קוולנטית )בצד 1Nשל הפירמידין או בצד 9Nשל הפורין( בקשר N-b- ל RNAלעומת זאת ,ידועים כמה תפקידים נוספים ,וכן ידועים כמה סוגים
glycosylלפחמן 1של הפנטוז ,והקבוצה הפוספטית מחוברת כאסטר של RNA. RNAריבוזומלי ) (rRNAהוא הרכיב המבני של הריבוזומים,
לפחמן 5של הפנטוז .הקשר N-b-glycosylנוצר על ידי דהידרציה )הסרת שתפקידם להוציא לפועל את סינתזת החלבונים בתא .מסנג'ר RNA
קבוצה קרבוקסילית מהפנטוז והסרת מימן מהבסיס החנקני( .כאשר ((mRNAהם מתווכים שנושאים מטען גנטי מגן או אחד או מכמה גנים אל
מולקולת הפוספט לא קיימת החומר הזה נקרא נוקלאוזיד ),(nucleoside הריבוזומים לצורך הסינתזה RNA .טרנספרלי ) (tRNAהם מולקולות
ובשם על פי בסיס חנקני מסויים זה יהיה :אדנוזין ,גאונוזין ,דאוקסיאדנוזין תומכות שמתרגמות את האינפורמציה של ה mRNAלרצף מסוים של
וכו'. חומצות אמינו .קיימים גם סוגים נוספים של RNAבעלי תפקידים ספציפיים.
בקצרה נעיר כי מלבד 5הבסיסים שהזכרנו שהם הנפוצים ביותר ב DNA בגלל הבדל משמעותי זה בין DNAו ,RNAהיחס בין דאוקסינוקלאוטידים
וב ,RNAישנם כמה בסיסים מינוריים שנפוצים הרבה פחות ,וככל הנראה )=של ה (DNAוהנוקלאוטידים הוא בין פי מאה לפי אלף לטובת
מטרתם היא שימור והגנה על המטען הגנטי .המספור והנומנקלטורה של הנוקלאוטידים; דאוקסי נוצרים רק בזמן שכפול של התא ,והנוקלאוטידים
הבסיסים הללו נעשית על ידי ציון השינוי מן הבסיס הרגיל )דוגמא :אם על עצמם נצרכים לכל אורך חיי התא.
פחמן 5של הבסיס המוכר ציטוזין נוספה קבוצת מתיל,
השם של הבסיס החדש הוא -5 נוקלאוטידים
.( methylcytosineבאותו אופן עשויים נוקלאוטידים הם בעלי שלשה מרכיבים מרכזיים:
להיות מספר קטן של נוקלאוטידים בהם בסיס חנקני )( 1
© נדב וילנר 1
יחידה - 8נוקלאוטידים בס"ד
ביוכימיים רבים בתא .המפורסם והנפוץ שבהם הוא ה הקבוצה הפוספטית אינה ממוקמת על פחמן 5של הפנטוז ,אלא על פחמן
ATPשהוא למעשה אדנזין-טרי-פוספט )שעכשיו אפשר 2או .3
להבין מה הוא ואיך הוא נראה( ,אבל גם נוקלאוסיידים
מרובי פוספטים אחרים משמשים כמטבעות אנרגיה הקשרים הפוספודיאסטרים
)למשל .(GTP הנוקלאוטידים משורשרים ביניהם ליצירת חומצות גרעין בעזרת קשר
שבירת הקשר האסטרי בין השרשרת הפוספטית והריבוז קוולנטי בין הקבוצה ההידרוקסילית על פחמן 3של הפנטוז עם הקבוצה
מניבה כ 14קילו-ג'אול ,ושבירת הקשר האנהידרידי בין הפוספטית שעל פחמן 5של הנוקלאוטיד הבא .קשר זה נקרא קשר
קבוצות הפוספט מניבה כ 30קילו-ג'אול ,ולדבר זה פוספודיאסטרי )כי זה פוספט בין שני אסטרים .(...יוצא שעמוד השדרה של
השפעה רבה על תהליכים תרמודינמיים המתרחשים ה DNAוה RNAהוא הידרופובי .ניתן לראות כי ב RNAנשארת הקבוצה
בתא. ההידרוקסילית על פחמן ,2וקבוצה זו יוצרת קשרי מימן עם מים .הקבוצה
הפוספטית בחומצות הגרעין טעונה ,כפי שניתן לראות ,שלילית ,ולכן
נוקלאוטידים משמשים כקופקטורים או רגולטורים תנוטרל בדרך כלל על ידי קשרים יוניים עם מטענים חיוביים על חלבונים,
האדנוזין משתתף רבות אנזימתיות בריאקציות יונים מתכתיים או פולימריים.
כקופקטור; הוא אינו משתתף ישירות בריאקציה כל הקשרים הפוספודיאסטריים בעלי אוריינטציה זהה בשרשרת ,ודבר זה
המרכזית ,אבל הסרתו של האדנוזין מהקופקטור עשויה נותן לחומצות הגרעין מבנה בעל פולריות ספציפית וקצה 5וקצה 3
לגרום לירידה בסדר גודל של מליונים בריאקטיביות של מוגדרים .בקצה 5לא יהיה נוקלאוטיד )אלא קבוצת פוספט( ובקצה 3לא
סובסטרט מסויים ,שכעת הקופקטור שלו לא מתפקד. תהיה קבוצה פוספט )אלא קבוצה הידרוקסילית(.
יתכן שהאדנוזין מסייע ל"המשכות" הסובסטרט אל האנזים ,ולאחר הסרתו באופן סכמטי ניתן לתאר את רצף הנוקלאוטידים בחומצות הגרעין כמתחיל
מהקופקטור הפעילות הזו מואטת באופן משמעותי .דוגמאות לדבר הם בקצה 5עם קבוצת פוספט המסומנת באות ,Pומסתיימת בקצה 3עם
קופקטורים חשובים כמו :קואנזים A, NADו ) FADשקשורים בשרשראות קבוצה קרבוקסילית .לכן ,תמיד מתארים חומצת גרעין מ : ` 3 `5מקצה
מעבר האלקטרונים ויצירת האנרגיה בתא(. 5לקצה ,3וניתן לכתוב זאת בכמה דרכים pACTGOH, pACTG :וכן הלאה.
מדוע דוקא אדנוזין? כנראה שהיות ובמהלך האבולציה הפך דוקא ה ATP חומצת גרעין של עד 50נוקלאוטידים תוגדר אוליגונוקלאוטיד ,ומעבר ל-
למטבע האנרגיה הבסיסי של התא ,נוח היה לבחור באדנוזין ,ששכיח 50תוגדר פולינוקלאוטיד.
ביותר בסביבה התאית ,לשמש כקופקטור ומשפיע על פעילות אנזימתית
בתא. תפקידים נוספים של נוקלאוטידים .2
ישנם נוקלאוטידים שמשמשים גם כמולקולות רגולטוריות .כך ,למשל,
נשיאת אנרגיה כימית בתאים
משמש cAMPשהוא וריאציה של אדנוזין כשליח משני בתא )=מעביר מידע
שנתקבל משליח ראשוני ,כמו הורמון ,אל תוך התא כדי להתחיל שרשרת הקבוצה הפוספטית עשויה לכלול במקרים מסויימים כמה קבוצות פוספט
פעולות תגובה בסביבה התוך תאית(. המשורשרות קוולנטית .השם ישתנה בהתאם :מונו ,די או טריפוספט וכן
הלאה ,כאשר מספור הקבוצות הוא אלפא ,בטא וכו' החל מהריבוז.
הידרוליזה של נוקלאוזיד טריפוספט מספקת אנרגיה המניעה תהליכים
© נדב וילנר 2
יחידה - 8נוקלאוטידים בס"ד
מגלוטמין לפחמן 1של ,PRPPשהוא ריבוזיל מטבוליזם של נוקלאוטידים .3
בעל 3קבוצות פוספט )ומולקולה חשובה
ישנם שני נתיבים מטבוליים ליצירת נוקלאוטידים:
ביותר בנושא שלנו – תרשים בהמשך( .שלב
זה מקוטלז ע"י גלוטמין-PRPP-אמידו-טרנספרז .התוצר של שלב פתיחה זה דה-נובו )(de novo pathway .1
הוא מאוד לא יציב ,ומכאן מתגלגל התהליך המטבולי ,כפי שמתואר בפירוט סלבג' )(salvage pathway .2
בספר .אנו יודעים כי אנזימים מסויימים מקטלזים אפילו כמה שלבים שונים סינתזת דה-נובו של נוקלאוטידים מתחילה בפריקורסורים )=חומרי
בשרשרת התהליכים. המוצא( המטבוליים שלהם :חומצות אמינו ,ריבוז-5-פוספט ,פחמן דו"ח ו 3N
חשוב לדעת כי רק בשלב 11של התהליך ישנו תוצר ביניים שמוגדר .Hסינתזת סלבג' למעשה ממחזרת את ה"שיירים" מפירוק של חומצות
פוריני והוא אינוזינט ,או .IMPהשלב הבא הוא שלב חשוב ,והסינתזה גרעין ונוקלאוסיידים.
מתפצלת לשתי דרכים אפשריות: סינתזת דה-נובו של נוקלאוטידים היא כנראה זהה בכל האורגניזימים.
.1יצירת – AMPע"י הוספת קבוצה אמינית הלקוחה הבסיסים החנקניים החופשיים )אדנין ,גואנין וכו'( אינם מתווכים בנתיב
מאספרטט ,ובשתי ריאקציות ,כאשר מולקולת GTP מטבולי זה; הם אינם מסונתזים ואז מוחברים אל הריבוז .למעשה ,הטבעת
משמשת כמקור אנרגטי לתהליך. הפורינית נבנת אטום אחר אטום ,ומחוברת אל הריבוז במהלך תהליך זה.
.2יצירת – GMPע"י חמצון בעזרת NAD+בפחמן 2של ה הטבעת הפירמידינית מסונתזת כ ) orotateטבעת( המחובר אל הפוספט ,ואז
,IMPוהשקעת אנרגיה ממולקולת .ATP עוברת המרה אל הטבעת הפירמידינת המוכרת .בסינתזת סלבג' הבסיסים
החופשיים לפעמים כן משמשים כמתווכים .כל המילים הגדולות האלה –
כדי שהדברים לא יהיו באוויר מצורף תרשים של המולקולות הנוצרות.
בפירוט בהמשך.
מנגנון הרגולציה של סינתזת דה-נובו של פורינים הוא ,feedback inhibition
ישנם כמה פריקורסורים חשובים בסינתזת דה-נובו של פירמידינים
כלומר ,התוצרי הסופיים AMPו GMPמווסתים את קצב והתקדמות
ופורינים PRPP .הוא פריקורסור חשוב ,וחומצות אמינו גם הן משחקות
התהליך .אמרנו קודם שהשלב הראשון בסינתזה הוא הצמדת חומצת אמינו
תפקיד מרכזי :גליצין כמקור לפורינים ואספרטט לפירמידין; גלוטמין
מגלוטמין ל .PRPPהאנזים שמקטלז שלב זה מעוכב הן על ידי IMPוהן ע"י
שמשמש ב 5 -שלבים שונים בסינתזה ,ואספרטט לפחות בשניים.
AMPו .GMPאותם תוצרים ממש מעכבים גם את הריאקציה בה מסונתז
הפחמנים בתהליך מתקבלים מטטרהידורפולאט )נרחיב עליו בהמשך(.
מגיב המוצא .PRPP
בשלב שני ,כאשר ישנו עודף גדול של GMPמעוכב האנזים ,xanthylate סינתזת דה-נובו של פורינים
ומסלול ) (2שבו נוצר GMPמעוכב ,בלי להשפיע על מסלול ) (1בו נוצר ה שני הנוקלאוטידים הפורינים המרכיבים את חומצות הגרעין הם אדנוזין-'5-
.AMPבאותו אופן מעכב עודף של AMPאת יצירתו ,בלא השפעה על .GMP מונופוספט ) (AMPוגואנוזין-'5-מונופוספט ) ,(GMPהמכילים את הבסיסים
מנגנון בקרה שלישי הוא עודף או חסר של ATPאו GTPבתא ,משום שכפי הפוריניים אדנין וגואנין .בשורות הבאות אנו הם התוצרים ואנו עוסקים
שראינו כדי ליצור AMPאנו זקוקים דוקא ל GTPואילו כדי ליצור GMPאנו בסינתזת דה-נובו שלהם ,כלומר ,בסינתזה שלהם ממטבוליטיים יסודיים
זקוקים למטבע האנרגיה המוכר .ATPכך שני המסלולים מבוקרים בנפרד. יותר.
את מקור החנקן והפחמן בטבעות הפוריניות ניתן לראות בתרשים .מבחינת
הסינתזה של הפורינים התהליך מתחיל בהצמדת חומצת אמינו הנתרמת
© נדב וילנר 3
יחידה - 8נוקלאוטידים בס"ד
מונופוספט .Vsתריפוספט סינתזת דה-נובו של פירמידינים
לאורך כל התיאור שחקנו כל הזמן בין AMPו ,ATPבין UMPו .UTPמה שני הנוקלאוטידים הפירמידיניים )לא להתבלבל עם פוריניים( המרכזיים
קורה כאן? בעצם מדובר על מספר הפוספטים המוצמדים לנוקלאוטיד, הם ציטידין-'5-מונופוספט ) (CMPויורידין-'5-מונופוספט ) .(UMPסינתזת
כאשר כדי להוסיף פוספט עושים פוספורילציה או תהליכים שמערבים דה-נובו שלהם שונה מזו של הפורינים ,משום שכאן אנו יוצרים קודם את
אנזימים גליקוליטיים .זו ריאקציה משותפת לכל התאיםATP + AMP : הטבעת הפירמידינית המשושה )כמעט לגמרי( ואז מחברים אליה את
2ADP במויל
קר ָ
הסוכר )ריבוז-5-פוספט( .לצורך התהליך המטבולי הזה משמש ָ
באותו אופן ישנם אנזימים שמבצעים פוספורילציה פוספט ) (carbomoyl phosphateכסובסטרט ראשוני ,והוא נוצר מגלוטמין
ATP + לנוקלאוזידים מונו או דו פוספטים .למשל: שמגיב עם ביקרבונט.
.NMP ADP + NDP קרבמויל פוספט מגיב עם אספרטט ,ויוצר את המולקולה שבתרשים .זהו
השלב הראשון המחייב בסינתזת הפירמידיניים ,והוא מסונתז על ידי אנזים
דאוקסיריבונוקלאוטידים מיוחד שחשוב לדעת את שמו – אספרטט טרנסקרבמוילז )או בשמוATCa :
דאוקסיריבונוקלאוטידים ,שהם אבני הבנין של ה ,DNA .seכלומר – מזרז ריאקציה בין אספרטט וקרבמויל( .תוצר השלב הזה הוא
מקורם בריבונוקלאוטידים מתאימים שעברו חיזור בפחמן 2 קרבמויל-אספרטט .לאחר מספר שלבים נוספים ודהידרציה מגיע שלב ה
שעל הריבוז ,ליצור את הצורה -'2דאוקסי )=בלא חמצן(. ,orotateכלומר ,נוצרת טבעת פירמידינית ,ואליה מוצמד הריבוז-5-פוספט
מבחינת הסימון נאמר כי ) ADPאדנוזין תרי פוספט( הופך שמקורו ב .PRPPבשלב זה נוצר ) UMPכלומר בעל פוספט אחד(,
ל =) dADPדאוקסיאדנוזין- לאחר החיזור ופוספורילציה יוצרת את ה UTPשהוא התוצר המבוקש .בעזרת אנזים
פוספט…( .האנזים המקטלז תרי- שנקרא cytidylate synthetaseהופך UTPל ,CTPובזה סיימנו לתאר את
הוא הזו הריאקציה את התהליך המטבולי.
) רידקטז ריבונוקלאוטיד רגולציה של הסינתזה נעשית ברובה דרך האנזים ) ATCaseשכזכור מקטלז
] (e reductaseרידקטז = מזרז ribonucleotid את השלב הראשון בתהליך( ,משום שהוא מעוכב על ידי התוצר .CTP
חיזור – נ.ו[. אנזים זה ,אצל בקטריות ,כולל 6תת -חידות קטליטיות ו 6תת-יחידות
החיזור לצורך רגולטוריות ,כאשר האיזור הקטליטי קושר את הסובסטרט והאיזור
אטומי נדרשים שני הרגולטורי את התוצר .CTPהאנזים קיים בשתי תצורות מבניות – פעיל ולא
מימן שמסופקים בשני מסלולים: פעיל; כאשר אין הרבה CTPוהאתרים הרגולטוריים חופשיים האנזים פעיל
שמקורם במולקולת NADPH מקסימלית .עם עלייה בקשירה של ה CTPאל האתרים הרגולטוריים
משתנה מבנה האנזים ,והאזורים הקטליטיים הופכים לא פעיליםATP .
בעזרת חלבון נושא מימן .1
מנטרל את השפעת ה ) CTPאם נחבר את זה לחומר קודם – CTPהוא
הנקרא .thioredoxinלחלבון זה יש
מודולטור שלילי ,ו ATPמודולטור חיובי(.
שני זוגות של קבוצות SHשמקבלות
את המימן מה ,NADPHריאקציה זו
מקוטלזת ע"י .thioredoxin reductase
© נדב וילנר 4
יחידה - 8נוקלאוטידים בס"ד
מסלול סלבג' ) (salvage pathway .4 חלבון נושא מימן אחר ,glutaredoxin -שגם הוא .2
כולל שני זוגות של ,SHכאשר מסלול זה מקוטלז ע"י
פורינים ופירמידיניים חופשיים משוחררים לתא לעתים תכופות ,בעיקר
.glutaredoxin reductase
במהלך דגרגציה של נוקלאוטידים .פורינים חופשיים בדרך כלל עוברים
"סלבג'" – כלומר" ,הצלה" ,ומנוצלים מחדש ליצירת נוקלאוטידים, פר"( מנוצליםס ֶ
הרעיון הוא שהחלבונים המחוזרים )=בעלי שני מימנים " ְ
במסלול מטבולי פשוט הרבה יותר מסינטזת דה-נובו. ע"י האנזים ריבונוקלאוטיד רידקטז והמימנים מועברים ל NDPליצירת
) dNDPשזה ניסוח כללי לכל נוקלאוטיד די-פוספט שהופך לדאוקסי(.
אחד ממסלול סלבג' המרכזיים הוא למעשה ריאקציה אחת המקוטלזת
בעזרת האנזים אדנוזין-פוספוריבוזיל-טרנספרז ,שבו אדנין חופשי מגיב עם האנזים ריבונוקלאוטיד רידקטז מהווה דוגמא למעורבות של רדיקלים
PRPPליצירת נוקלאוטיד אדנין ,על פי הריאקציהAdenine + PRPP : חופשיים במכניזם של הריאקציה .לאנזים שתי תת יחידות ,R1, R2שכל
.AMP + PPi אחת מחולקת לשתיים 1R .כוללת שני אתרים רגולטוריים ,והאתרים
הפעילים )המקטלזים( נמצאים בחיבור בין 1Rל .2Rקשירה של רדיקלים
חופשיים של ברזל ליחידה 2Rגורמת שינויים באתר הפעיל ומאפשרת את
פעולת הזירוז של האנזים .אנחנו מכירים 4סוגים של האנזים
ריבונוקלאוטיד רידקטז ,כאשר כל סוג נבדל מחברו בקבוצה המספקת את
הרדיקל החופשי ובקופקטור הדרוש ליצירתו.
נקודה נוספת וחשובה לגבי האנזים קשורה בכך שיש לו ,כפי שציינו ,שני
אתרים רגולטוריים .אתר אחד מאקטב או מנטרל את פעילות האנזים
בהתאם לקשירה ל ) ATPמודולטור חיובי( או לתוצר ) dNDPמודולטור
שלילי( .האתר השני מסוגל לשנות את הספציפיות שלו לפי המולקולות
- dCDPתכונה ייחודית זו של הנקשרות אליו .עודף dATPיגרום ליצירת
האנזים מאפשרת אספקה מאוזנת ביותר בין פורינים ופירדמידינים לצורך
יצירת ה .DNA
פירוק של פורינים ופירמידינים
נוקלאוטידים פורינים עוברים דגרגציה בנתיב מטבולי שהוא למעשה הפוך
לנתיב סינתזת דה-נובו שלהם :הם מאבדים פוספט ,הופכים לאדנוזין
מאבדים קבוצה אמינית והופכים לאינוזין עוברים הידרוליזה ואז
רית ) .(uric acidקטבוליזם של
אוקסידציה והופכים ל xanthineוחומצה או ִ
GMPהוא תהליך דומה ,וגם בסופו נוצרת חומצה אורית .החומצה נוצרת או
מקטבוליזם של פורינים או מיצירת חומצות גרעין ומופרשת מהגוף ) 0.6ג'
ל 24-שעות( .דגרגציה של נוקלאוטידים פירמידיניים מובילה בדרך כלל
ליצירת ,+NH4וממילא לסינתזה של אוריאה.
© נדב וילנר 5
יחידה - 8נוקלאוטידים בס"ד
גם פורינים אחרים יכולים לעבור ריאקצית סלבג' דומה; האנזים שמקטלז
את מסלול הסלבג' של גואנין נקרא ,HGPRTוגם לפירמידינים קיים מסלול
מטבולי כזה )אבל בעיקר אצל חיידקים ובע"ח יונקים – לא באדם!( .חסר
באנזים שמזרז ריאקציה זו עבור גואנין בא לידי ביטוי בתסמונת ייחודית
שנקרא לש-נייהן ) ;(Lesch-Nyhanשסימניה הם פיגור קשה ובעיות
Amino acids מוטוריות .דבר זה מלמד אותנו על המשמעות הגדולה של מסלול הסלבג'.
ATP במידה והאנזים הנ"ל חסר ,רמות PRPPבתא עולות ויש יותר סינתזת דה-
Ribose-5-P נובו של פורינים ,דבר שגורם לרמות גבוהות של חומצה אורית בתא ,דבר
Folic acid שעשוי לגרום למחלת .Goutהמסלולים המטבוליים המתוארים תלויים
ומשפיעים זה על זה.
מחלת Goutואנטימטבוליטים בסינתזת נוקלאוטידים .5
Gout
IMP Nucleotidesוממילא את
את יצירת ה שבעקיפין מעכב
in diet כפי שראינו לעיל ,ייצור מוגבר של חומצה אורית גורם למחלת gout
התהליך כולו. )בתמונה( .למחלה זו יש השפעה על הפרקים ,אבל שקיעה של חומצה
אורית מתרחשת גם בטיובולים של הכלייה; המחלה נפוצה בעיקר בגברים,
והאטיולוגיה שלה אינה ידועה לגמרי ,אך נחשבת גנטית.
הטיפול במחלה הוא קודם כל ע"י אכילת מזון דל בנוקלאוטידים ,כדי למנוע
פעילות יתר של מסלול הסלבג' .דרך נוספת היא תרופתית ,והיא מתבססת
על אלופירינול ) ,(Allopurinolשתפקידו בעצם לעכב את הקסנטין-אוקסידז
שמקטלז את הריאקציה שבה הופכים פורינים )היפוקסנטין( לחומצה
אנטימטבוליטים אחרים אורית .הדבר גורם לכך שבמקום לפלוט חומצה אורית ,הגוף פולט קסנטין
במחלות סרטניות ,באופן הגיוני ,ישנה והיפוקסנטין ,שהם תוצרים עם נטייה נמוכה ביותר להפוך לגבישים
פעילות סנתוז מוגברת של .DNAלכן, )בהשוואה לחומצה אורית(.
הקונספט של עיכוב המטבוליזם והסינתזה האלופורינול פועל בשתי דרכים מקבילות:
של הנוקלאוטידים הוא אחד מן הבסיסיים תחרות עם היפוקסנטין – בגלל דמיונו של האלופורינול .1
בחיפוש אחר תרופה למחלות ממאירות. למולקולת ההיפוקסנטין ,כאשר הוא נפגש עם האנזים הוא הופך
דוגמאות לאנטימטבוליטים שכאלה: לאוקסיפורינול ,ונקשר אל האתר הפעיל של האנזים ומעכב אותו.
– azaserine, acivicin .1שתי מולקולות אלה דומות במיוחד לגלוטמין, צריכת – PRPPהאלופורינול מקוטלז גם על ידי HGPRT .2
ולכן משמשות כמעכב יעיל של האנזים גלוטמין-אמידו-טרנספרז לאלופורינול-ריבוטייד ,ובריאקציה זו הוא צורך ,PRPPדבר
© נדב וילנר 6
יחידה - 8נוקלאוטידים בס"ד
מסלול הסלבג' .היות וידוע כי PRPPהוא רגולטור בשלב המחייב בסינתזת )שתפקידו כפי שראינו לתרום את הקבוצה החנקנית בתהליכי סינתזה
דה-נובו של פורינים ,עלה החשד כי ה PRPPסינתטז פגום בתכונות רבים של נוקלאוטידים(.
הרגולציה שלו ,אך לא בתכונותיו הקטליטיות. – fluorouracil .2הופך בתא במסלול סלבג' ל FdUMPשנקשר
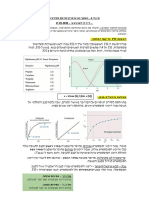
בניסוי נבדקה פעילות האנזים בתנאים שונים – לא רק פיזיולוגיים, קוולנטית – ובעצם עושה דהאקטיבציה – לאנזים תימידילט-סינתטז,
ובריכוזים קבועים של ,ADP, R-5-P שהכרחי במסלול המטבולי היחיד הקיים בתא ליצירת תימין!
ופוספט אנאורגני .נמצא כי בהעלאת – methotrexate, aminopterin .3מעכבים את האנזים dihydrofolate
ריכוז הפוספט האנאורגני אנזים reductaseשהוא האנזים שמחזר את החומצה הפולית )שעליה נרחיב
נורמלי מדגים גרף סיגמואידי בהמשך( ,והופך אותה למולקולה האקטיבית טטרהידרופולאט.
ואילו האנזים ,ADP בנוכחות האנטימטבוליטים הללו מתפקדים כמעכבים תחרותיים.
המוטנטי מדגים גרף היפרבולי.
כאשר נוקו מן המערכת כל מולקולות מרכזיות בסינתזת נוקלאוטידים .6
המעכבים של האנזים – שני
האנזימים הפגינו תגובה היפרבולית PRPP סינתזת
)ואפילו נמצא כי האנזים המוטנטי כדי להשלים את החלק הזה של הדיון נזכיר את הריאקציה שבה מייצרים
פחות רגיש למעכבים(. את מולקולת ה PRPPשהיא מולקולה מרכזית בסינתזה של נוקלאוטידים.
המסקנה היתה כי בריכוז פיזיולוגי של פוספט אנאורגני האנזים הנורמלי נוסחת הריאקציה היא:
הופך רגיש למעכבים תאיים שונים ,ואילו האנזים המוטנטי פגום בדיוק Ribose-5-phosphate + ATP 5-phosphoribosyl-1-pyrophosphate + AMP
בתכונה זו .בשילוב עם העובדה שפעילותו הקטליטית בתנאים לא וצורת המולקולה עצמה:
פיזיולוגיים הוכחה כזהה לזה של האנזים הנורמלי ניתן להסיק כי האנזים
הוא אלוסטרי ,והפגם שלו הוא באתר הרגולטורי ,שאמור לאקטב את
מסלול הדה-נובו ,כפי שצויין לעיל.
חומצה פולית וטטרהידרופולאט
חומצה פולית הכרחית לקיום מסלולי מטבוליזם רבים בגוף ,בין השאר
סינטזה של נוקלאוטידים .בני אדם מקבלים את החומצה הפולית הדרושה הריאקציה הזו מזורזת ע"י ,PRPP synthetaseומתרחשת בנוכחות מגנזיום
להם מן המזון ,ואילו חיידקים מסנטזים את החומצה לעצמם .תכונה זו ניתן ופוספט אנאורגני .במאמר ) (Sperling, Medicine, 1973מתואר ניסוי שנעשה
לנצל כדי לייצר חומרים אנטיבקטריאליים ,כגון סולפה )סולפנילאמיד(, במוטציה של האנזים PRPPסינתטז .המקרה הוא של חולה זכר בן 35
שמעכבים את הסינטזה של הח' הפולית אצל הבקטריות ,אך לא משפיעים שסובל כ 15 -שנה מ ) goutצינית( ,רמות החומצה האורית בדמו גבוהות
באופן ציטוטוקסי על יונקים ובני אדם .מבנה החומצה: מאוד בהשוואה לנורמל ,ופעילות האנזימים היפוקסנטין ו HGPRTתקינה.
כפי שראינו ,פעילות יתר של מסלול הסלבג' עשויה לגרום לצינית ,ומכאן
שכנראה יש לחולה בעיה מסלול הדה-נובו ,ולכן משופעל אצלו באופן חריג
© נדב וילנר 7
יחידה - 8נוקלאוטידים בס"ד
הסולפה דומה במבנהו לאמינו-בנזואט שבמרכז המולקולה ,ולכן מעכב את
הסינתזה .חסר בחומצה פולית הוא מחלת חסר ויטמין הנפוצה ביותר ,והוא
גורם – בתור התחלה – לאנמיה מגלובלסטית ,כלומר ,לבעייה בייצור
אריתרוציטים תקינים .סיבת הדבר ,כפי שנראה ,היא שהחומצה היא מצרך
בסיסי בסינתזת נוקלאוטידים ,ולכן בתאים המתחלקים הרבה תווצר בעיה –
במקרה שלנו ,בהמופואזיס שבמח העצם.
אחד הקופקטורים המשמעותיים במטבוליזם של נוקלאוטידים הוא
טטרהידרופולאט ) .(tetrahydrofolateטטרהידרופולאט )(H4 folate or THF
מורכב למעשה מקבוצה פטרית ,מאמינובנזן ומגלוטמט ,והוא תוצר חיזור
של חומצה פולית:
את החיזור מבצע אנזים שנקרא ,dihydrofolate reductaseשמשתמש ב
NADPHכסוכן חיזור .כפי שהזכרנו בסעיף הקודם ישנם אנטימטבוליטים
הצורה השמאלית ביותר של THFמעורבת ביצירת חומצת האמינו מתיונין
שדומים לח' פולית ,ומשמשים כמעכבים תרפוייטים במקרים של סרטן,
בכבד .בריאקציה זו הופך הומוציסטאין למתיונין ,בסיוע methyl-THF-5
משום שהוא מעכבים את היצירה של .THF
ובסיוע הויטמין .12Bהאנזים שמקטלז את הריאקציה זקוק לויטמין ,12B
THFיכול לעבור בין מצבי חיזור וחמצון שונים ,בהתאם למצב החנקן
ולמעשה הוא האנזים היחיד שהופך את ה methyl-THF-5ל ,THFכאשר
במקום 5ובמקום :10אם מחובר לפחמן 5קבוצת מתיל זהו המצב המחוזר
אנו יודעים שה THFנצרך לריאקציות רבות נוספות בגוף .משמעות הדבר
ביותר ,אם מחובר אליו פחמן דו"ח זה מצב ביניים ,ואם מחובר אליו פורמיל
שחסר ב ,12Bיגרום להצברות של methyl-THF-5בדם וברקמות ולחסר ב
– זהו המצב המחוזר ביותר .באופן זה יכול טטרהידרופולאט לשמש
,THFובסופו של דבר לאנמיה ואולי להפרעות קליניות נוספות.
כ"תורם" קבוצות פחמן-יחיד )כמו מתיל( בריאקציות שונות בתא .מצבי
חיזור-חמצון של ) THFהמחומצן ביותר בימין(:
© נדב וילנר 8
יחידה - 8נוקלאוטידים בס"ד
© נדב וילנר 9
You might also like
- סיכימים ביוכימיה בDocument72 pagesסיכימים ביוכימיה בapi-3717496100% (2)
- חוברת אנטומיהDocument392 pagesחוברת אנטומיהGaia Shcherbina100% (1)
- תרגול 10Document47 pagesתרגול 10Shahar Tam100% (1)
- וכימה כללית וקלינית-אנזימיםDocument23 pagesוכימה כללית וקלינית-אנזימיםapi-25967322100% (1)
- סיכום לביוכימיה א שיר אורנשטיין 1Document60 pagesסיכום לביוכימיה א שיר אורנשטיין 1בן הרושNo ratings yet
- ביוכימיה5Document8 pagesביוכימיה5Zohar ShaharabaniNo ratings yet
- ביוכימיה5Document8 pagesביוכימיה5Zohar ShaharabaniNo ratings yet
- סכום הסטולוגיה2012Document36 pagesסכום הסטולוגיה2012Jasmin KolodroNo ratings yet
- ביולוגיה של התא-לימודי תואר ראשון של מדעי החיים- סופר סיכוםDocument41 pagesביולוגיה של התא-לימודי תואר ראשון של מדעי החיים- סופר סיכוםyuliakesler83% (6)
- חומר טוב ביולוגיה של התאDocument50 pagesחומר טוב ביולוגיה של התאDaniel Shtriker67% (3)
- סיכום משופר ביוכימיהDocument34 pagesסיכום משופר ביוכימיהrichter1No ratings yet
- ביולוגיה מולקולרית סיכוםDocument75 pagesביולוגיה מולקולרית סיכוםAnat Lyubin Haimov100% (2)
- ביוכימיה4Document4 pagesביוכימיה4Zohar ShaharabaniNo ratings yet
- סיכום גנטיקהDocument52 pagesסיכום גנטיקהilana shelegNo ratings yet
- תקשורת בין נוירוניםDocument6 pagesתקשורת בין נוירוניםYiska AmarNo ratings yet
- סיכום לקורס ביולוגיה מולקולרית וביטוי גנים תשסגDocument31 pagesסיכום לקורס ביולוגיה מולקולרית וביטוי גנים תשסגapi-3824802100% (2)
- שכפול דנא ומ-דנא לחלבוןDocument12 pagesשכפול דנא ומ-דנא לחלבוןak_mrNo ratings yet
- ביוכימיה א חלק א דרורDocument31 pagesביוכימיה א חלק א דרורGal ZeevNo ratings yet
- Biology AnswersDocument5 pagesBiology Answershagaraa15No ratings yet
- פרק 7 - פוטוסינתזהDocument4 pagesפרק 7 - פוטוסינתזהyuliakeslerNo ratings yet
- מצגת מעודכן 8Document22 pagesמצגת מעודכן 8AseelNo ratings yet
- מושגים כחולים במיקרוביולוגיהDocument9 pagesמושגים כחולים במיקרוביולוגיהliaorNo ratings yet
- ביולוגיה של התא סיכום אינטרט 4Document82 pagesביולוגיה של התא סיכום אינטרט 4Emily PeretzNo ratings yet
- נוסחאון ביוכימיה 2Document2 pagesנוסחאון ביוכימיה 2api-3806097No ratings yet
- שיעור 2 - אריזת הדנא בגרעיןDocument4 pagesשיעור 2 - אריזת הדנא בגרעיןShirel YehezkelNo ratings yet
- מושגים בביולוגיהDocument29 pagesמושגים בביולוגיהdiegomano100% (1)
- סיכומים להרצאות 1 5Document10 pagesסיכומים להרצאות 1 5annaerelNo ratings yet
- דף ביוכימיה-שירליDocument9 pagesדף ביוכימיה-שירליAlaa HajNo ratings yet
- שעתוקDocument7 pagesשעתוקapi-19979047No ratings yet
- שיעור 6 - המשך אנזימים וקינטיקת סטורציהDocument12 pagesשיעור 6 - המשך אנזימים וקינטיקת סטורציהtamar.razNo ratings yet
- ביוכימיה א חלק ב דרורDocument35 pagesביוכימיה א חלק ב דרורGal ZeevNo ratings yet
- סיכום במדעי הצמחDocument59 pagesסיכום במדעי הצמחEinav Stern100% (1)
- סיכום גן לחלבוןDocument19 pagesסיכום גן לחלבוןapi-3831508No ratings yet
- תרגוםDocument6 pagesתרגוםapi-19979047No ratings yet
- סיכום פרק מדנא לחלבוןDocument9 pagesסיכום פרק מדנא לחלבוןxliavx11No ratings yet
- Bio Chem Formula So LaDocument4 pagesBio Chem Formula So Laapi-3806097No ratings yet
- סיכום למבחןDocument13 pagesסיכום למבחןnoanevo11No ratings yet
- Lesson15 30012008Document8 pagesLesson15 30012008api-25967322100% (2)
- נועה רוזנטל- שיטות מחקר בביולוגיה מולקולריתDocument26 pagesנועה רוזנטל- שיטות מחקר בביולוגיה מולקולריתZiv ItzkovichNo ratings yet
- מיקרוביולוגיה - שיעור 2 - 050308Document9 pagesמיקרוביולוגיה - שיעור 2 - 050308api-3738386100% (1)
- גנטיקה - שיעור 17 - 231207Document6 pagesגנטיקה - שיעור 17 - 231207api-3738386No ratings yet
- ביוכימיה א-שליף דף1Document1 pageביוכימיה א-שליף דף1api-3831508100% (1)
- ביוכימיה א' דף שליף 3Document3 pagesביוכימיה א' דף שליף 3api-3831508100% (1)
- 20Document3 pages20netaa6No ratings yet
- שיעור 10 - מבוא לסוכריםDocument8 pagesשיעור 10 - מבוא לסוכריםtamar.razNo ratings yet
- מעגל קלוויןDocument17 pagesמעגל קלוויןliaorNo ratings yet
- שיטות סמסטר ב סיכום1Document34 pagesשיטות סמסטר ב סיכום1Nitzan ArgamanNo ratings yet
- וירוסים מחברת תשפאDocument46 pagesוירוסים מחברת תשפאShir DahanNo ratings yet
- מיטוכונדריהDocument4 pagesמיטוכונדריהapi-19979047No ratings yet
- פרק 3 - התא הצמחיDocument5 pagesפרק 3 - התא הצמחיyuliakeslerNo ratings yet
- 12 פוטוסינתזהDocument7 pages12 פוטוסינתזהירדן לויןNo ratings yet
- תשובות למעבדה 1Document1 pageתשובות למעבדה 1Shira GoldbergNo ratings yet
- 1 - חומצות אמינוDocument4 pages1 - חומצות אמינוtamar.razNo ratings yet
- מיקרוביולוגיה - שיעור 3 - 120308Document11 pagesמיקרוביולוגיה - שיעור 3 - 120308api-3738386100% (1)
- 6 סינתזת חלבוןDocument18 pages6 סינתזת חלבוןMatanAlayofNo ratings yet
- ביולוגיה של התא - זרימת המידע בתאDocument6 pagesביולוגיה של התא - זרימת המידע בתאGal ZeevNo ratings yet
- +שוליים+לינה-דף 2Document2 pages+שוליים+לינה-דף 2api-3806097No ratings yet
- אנטומיה ופיזיולוגיה לסיעוד - רמת ארגון של התא וסוגי הרקמות - 14984611Document37 pagesאנטומיה ופיזיולוגיה לסיעוד - רמת ארגון של התא וסוגי הרקמות - 14984611safa titiNo ratings yet
- דגם תאDocument10 pagesדגם תאHaim EzerNo ratings yet
- מגן לחלבון תשפב - שיעור 1Document22 pagesמגן לחלבון תשפב - שיעור 1saja MuhammadNo ratings yet
- ביוכימיה3Document4 pagesביוכימיה3Zohar ShaharabaniNo ratings yet
- יחידה8Document9 pagesיחידה8Zohar ShaharabaniNo ratings yet