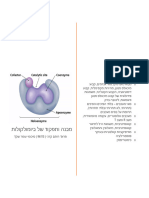
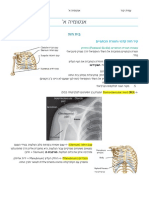
You might also like
- - - סיכום אנטומיה - עמית מנור - בטן - מעודכן 270320Document17 pages- - סיכום אנטומיה - עמית מנור - בטן - מעודכן 270320tamar.raz100% (1)
- סיכום יערה- ביולוגיה של התאDocument83 pagesסיכום יערה- ביולוגיה של התאtamar.razNo ratings yet
- סיכום משופר ביוכימיהDocument34 pagesסיכום משופר ביוכימיהrichter1No ratings yet
- אורגנית סיכומוןDocument8 pagesאורגנית סיכומוןRaphael IsraeliNo ratings yet
- עזוב אותי באימש'ך-דוח 4Document21 pagesעזוב אותי באימש'ך-דוח 4api-3801666No ratings yet
- ביוכימיה5Document8 pagesביוכימיה5Zohar ShaharabaniNo ratings yet
- סיכימים ביוכימיה בDocument72 pagesסיכימים ביוכימיה בapi-3717496100% (2)
- סיכום תגובות בכימיה אורגניתDocument11 pagesסיכום תגובות בכימיה אורגניתEliran SabagNo ratings yet
- שיעור 3- המשך שיטות הפרדת חלבונים ומבנה תלת מיימדיDocument16 pagesשיעור 3- המשך שיטות הפרדת חלבונים ומבנה תלת מיימדיtamar.razNo ratings yet
- סיכומים קרדיולוגיהDocument9 pagesסיכומים קרדיולוגיהarielNo ratings yet
- מבנה האטום (מצגת 7)Document8 pagesמבנה האטום (מצגת 7)tamar.razNo ratings yet
- בית חזה - עמית יבורDocument75 pagesבית חזה - עמית יבורtamar.razNo ratings yet
- וכימה כללית וקלינית-אנזימיםDocument23 pagesוכימה כללית וקלינית-אנזימיםapi-25967322100% (1)
- מצגת 6 שריריםDocument26 pagesמצגת 6 שריריםShahar TamNo ratings yet
- סיכומים בביוכימיהDocument3 pagesסיכומים בביוכימיהapi-3717496No ratings yet
- סיכום אנזימיםDocument3 pagesסיכום אנזימיםrachel112100% (1)
- דו'ח 5 - מוליכות ופוטנציומטריה גיאDocument9 pagesדו'ח 5 - מוליכות ופוטנציומטריה גיאLeon DobrzinskyNo ratings yet
- מרתון כימיהDocument6 pagesמרתון כימיהS.N.HNo ratings yet
- ביוכימיה - עד ליפידיםDocument149 pagesביוכימיה - עד ליפידיםtamar.razNo ratings yet
- סיכומון ביוכימיה ללא קרישה 2025Document84 pagesסיכומון ביוכימיה ללא קרישה 2025tamar.raz100% (2)
- תקשורת בין נוירוניםDocument6 pagesתקשורת בין נוירוניםYiska AmarNo ratings yet
- תרגול 10Document47 pagesתרגול 10Shahar Tam100% (1)
- כוחות בין מולקולריים (מצגת 9)Document7 pagesכוחות בין מולקולריים (מצגת 9)tamar.raz100% (1)
- דף למבחן ביוכימיהDocument3 pagesדף למבחן ביוכימיהliaorNo ratings yet
- אנזימים תשפבDocument63 pagesאנזימים תשפבNicole FrumkinNo ratings yet
- Enzymes 2022Document64 pagesEnzymes 2022mayah sabarNo ratings yet
- ביולוגיה של התא סיכום אינטרט 4Document82 pagesביולוגיה של התא סיכום אינטרט 4Emily PeretzNo ratings yet
- שליף מעודכןDocument4 pagesשליף מעודכןGalia KlarNo ratings yet
- 3Document7 pages3Dana ShoganNo ratings yet
- מושגים כחולים במיקרוביולוגיהDocument9 pagesמושגים כחולים במיקרוביולוגיהliaorNo ratings yet
- ביולוגיה - אנרגיה וריאקציותDocument2 pagesביולוגיה - אנרגיה וריאקציותroeiamediNo ratings yet
- שיעור 16 - קומפלקס פירובט דהידרוגנאז ומעגל קרבסDocument14 pagesשיעור 16 - קומפלקס פירובט דהידרוגנאז ומעגל קרבסtamar.razNo ratings yet
- 4- אנזימיםDocument16 pages4- אנזימיםtamar.razNo ratings yet
- יחידה8Document9 pagesיחידה8Zohar ShaharabaniNo ratings yet
- ביוכימיה א חלק א דרורDocument31 pagesביוכימיה א חלק א דרורGal ZeevNo ratings yet
- נוסחאון ביוכימיה 2Document2 pagesנוסחאון ביוכימיה 2api-3806097No ratings yet
- ביוכימיה א-שליף דף1Document1 pageביוכימיה א-שליף דף1api-3831508100% (1)
- סיכום מבנה ותפקוד של ביומולקולות - פרופ רותם קרניDocument46 pagesסיכום מבנה ותפקוד של ביומולקולות - פרופ רותם קרניMeitar AtiasNo ratings yet
- פרק 7 - פוטוסינתזהDocument4 pagesפרק 7 - פוטוסינתזהyuliakeslerNo ratings yet
- מיטוכונדריהDocument4 pagesמיטוכונדריהapi-19979047No ratings yet
- שיעור 10 - מבוא לסוכריםDocument8 pagesשיעור 10 - מבוא לסוכריםtamar.razNo ratings yet
- מושגים בביולוגיהDocument29 pagesמושגים בביולוגיהdiegomano100% (1)
- פרק 12 אבלינהDocument19 pagesפרק 12 אבלינהG GNo ratings yet
- Bio Chem Formula So LaDocument4 pagesBio Chem Formula So Laapi-3806097No ratings yet
- Protocol Enzymology Lab תשפדDocument8 pagesProtocol Enzymology Lab תשפדolir443No ratings yet
- מעבדהDocument13 pagesמעבדהEmad AwadNo ratings yet
- פיזיולוגיה של מערכות - שיעור 4 - 200308Document9 pagesפיזיולוגיה של מערכות - שיעור 4 - 200308api-3738386No ratings yet
- ביוכימיה א חלק ג דרורDocument72 pagesביוכימיה א חלק ג דרורGal ZeevNo ratings yet
- שיעור 7 19.3Document2 pagesשיעור 7 19.3tamar.razNo ratings yet
- שיטות סמסטר ב סיכום1Document34 pagesשיטות סמסטר ב סיכום1Nitzan ArgamanNo ratings yet
- - - סיכום באנטומיה ופיזיולוגיה -Document57 pages- - סיכום באנטומיה ופיזיולוגיה -Arkan EssaNo ratings yet
- מצגת מעודכן 8Document22 pagesמצגת מעודכן 8AseelNo ratings yet
- ביולוגיה 4Document2 pagesביולוגיה 4Eran ReznikNo ratings yet
- דף ביוכימיה-שירליDocument9 pagesדף ביוכימיה-שירליAlaa HajNo ratings yet
- דף נוסחאות ביוכימיה ב'Document3 pagesדף נוסחאות ביוכימיה ב'liaorNo ratings yet
- 1 - חומצות אמינוDocument4 pages1 - חומצות אמינוtamar.razNo ratings yet
- Chapter 17 - Carbonilic IDocument12 pagesChapter 17 - Carbonilic IahmaszitimNo ratings yet
- תרגיל 2-מטבוליזם וחילוף החומריםDocument3 pagesתרגיל 2-מטבוליזם וחילוף החומריםsaja MuhammadNo ratings yet
- דף נוסחאות גיאוכימיהDocument2 pagesדף נוסחאות גיאוכימיהTomer GadolNo ratings yet
- 1 144938779Document62 pages1 144938779Svetlana AzizovNo ratings yet
- אלקטרו פיזיולוגיה אסף סיכוםDocument11 pagesאלקטרו פיזיולוגיה אסף סיכוםאייל אברהםNo ratings yet
- וירוסים מחברת תשפאDocument46 pagesוירוסים מחברת תשפאShir DahanNo ratings yet
- Copy of סיכום ביומולקולות 2016 - דניאל ויקטורDocument60 pagesCopy of סיכום ביומולקולות 2016 - דניאל ויקטורMeitar AtiasNo ratings yet
- 8 גלוקונאוגנזהDocument6 pages8 גלוקונאוגנזהירדן לויןNo ratings yet
- 13 מטבוליזם של חומצות אמינוDocument7 pages13 מטבוליזם של חומצות אמינוירדן לויןNo ratings yet
- ביוכימיה א' דף שליף 3Document3 pagesביוכימיה א' דף שליף 3api-3831508100% (1)
- למידה עצמית עצבוב בית החזה, ניקוז לימפטיDocument11 pagesלמידה עצמית עצבוב בית החזה, ניקוז לימפטיtamar.razNo ratings yet
- שיעור 7 19.3Document2 pagesשיעור 7 19.3tamar.razNo ratings yet
- ביוסינתזה של חומצות אמינו 2.4.20Document3 pagesביוסינתזה של חומצות אמינו 2.4.20tamar.razNo ratings yet
- בית חזהDocument73 pagesבית חזהtamar.razNo ratings yet
- שיעור 21 - המשך חימצון חומצות שומות, גופי קטון וביוסניתזה של חומצות שומןDocument10 pagesשיעור 21 - המשך חימצון חומצות שומות, גופי קטון וביוסניתזה של חומצות שומןtamar.razNo ratings yet
- מנגנוני טרנספורטDocument39 pagesמנגנוני טרנספורטtamar.razNo ratings yet
- 3 - מבנה החלבוןDocument15 pages3 - מבנה החלבוןtamar.razNo ratings yet
- ביולוגיה של התא- לוטם קוןDocument175 pagesביולוגיה של התא- לוטם קוןtamar.raz100% (1)
- שיעור 1- מבוא - התא- שיטות מחקרDocument2 pagesשיעור 1- מבוא - התא- שיטות מחקרtamar.razNo ratings yet
- שיעור 9 - ביואנרגיהDocument12 pagesשיעור 9 - ביואנרגיהtamar.razNo ratings yet
- 4- אנזימיםDocument16 pages4- אנזימיםtamar.razNo ratings yet
- שיעור 5Document2 pagesשיעור 5tamar.razNo ratings yet
- 1 - חומצות אמינוDocument4 pages1 - חומצות אמינוtamar.razNo ratings yet
- שיעור 10 - מבוא לסוכריםDocument8 pagesשיעור 10 - מבוא לסוכריםtamar.razNo ratings yet
- 2 - שיטות ניתוח חלבוניםDocument10 pages2 - שיטות ניתוח חלבוניםtamar.razNo ratings yet
- ביופיזיקה - מאוחד (אופיר)Document75 pagesביופיזיקה - מאוחד (אופיר)tamar.razNo ratings yet
- גזים (מצגת 5)Document5 pagesגזים (מצגת 5)tamar.raz100% (1)
- בית חזה - הילה מאי - שחר דקלDocument79 pagesבית חזה - הילה מאי - שחר דקלtamar.razNo ratings yet
- שיעור 16 - קומפלקס פירובט דהידרוגנאז ומעגל קרבסDocument14 pagesשיעור 16 - קומפלקס פירובט דהידרוגנאז ומעגל קרבסtamar.razNo ratings yet
- שיעור 11- קליטת סוכרים במעי וטרנספורטרים לגלוקוזDocument6 pagesשיעור 11- קליטת סוכרים במעי וטרנספורטרים לגלוקוזtamar.razNo ratings yet
- סיכום ביופיזיקה שיר ולבר וטל קולרDocument150 pagesסיכום ביופיזיקה שיר ולבר וטל קולרtamar.razNo ratings yet
- שיעור 2 אפידימולוגיה מדדי תחלואה ותמותה אמבר 4.11.19Document6 pagesשיעור 2 אפידימולוגיה מדדי תחלואה ותמותה אמבר 4.11.19tamar.razNo ratings yet