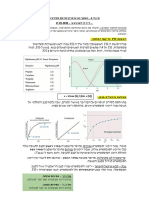
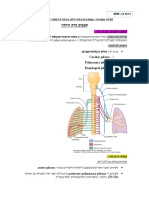
You might also like
- ביוכימיה4Document4 pagesביוכימיה4Zohar ShaharabaniNo ratings yet
- מעבדהDocument13 pagesמעבדהEmad AwadNo ratings yet
- שיעור 3- המשך שיטות הפרדת חלבונים ומבנה תלת מיימדיDocument16 pagesשיעור 3- המשך שיטות הפרדת חלבונים ומבנה תלת מיימדיtamar.razNo ratings yet
- הגשה 2Document6 pagesהגשה 2peterNo ratings yet
- ביולוגיה סיכוםDocument56 pagesביולוגיה סיכוםמירי צבריNo ratings yet
- 232254469Document40 pages232254469asherbigNo ratings yet
- מעבדה אחרונה וסופיתDocument5 pagesמעבדה אחרונה וסופיתapi-3801666No ratings yet
- דף ביוכימיה-שירליDocument9 pagesדף ביוכימיה-שירליAlaa HajNo ratings yet
- NoName 04052020071546Document5 pagesNoName 04052020071546Bayan HamedNo ratings yet
- דוח מסכם אסתריפיקציהDocument16 pagesדוח מסכם אסתריפיקציהDaniel Turgeman HaniaNo ratings yet
- דף עזר שחרDocument15 pagesדף עזר שחרShahar TamNo ratings yet
- Lab Report בביוכימיה מס' 1Document4 pagesLab Report בביוכימיה מס' 1k2703kNo ratings yet
- דוח מכין טרימיריסטיןDocument5 pagesדוח מכין טרימיריסטיןאנונימיNo ratings yet
- סיכומי מצעיםDocument3 pagesסיכומי מצעיםgelb50% (2)
- דוח מעבדה בהנדסה גנטית - שיבוט וטרנספורמציהDocument10 pagesדוח מעבדה בהנדסה גנטית - שיבוט וטרנספורמציהAmir Erel100% (1)
- ניקוי ליזוזיםDocument6 pagesניקוי ליזוזיםliaorNo ratings yet
- סיכום אנזימיםDocument3 pagesסיכום אנזימיםrachel112100% (1)
- מושגים בביולוגיה של התא - 1Document11 pagesמושגים בביולוגיה של התא - 1Jasmin KolodroNo ratings yet
- מצגת 8 ביוכימיה יסודות ואנזימים 2022Document115 pagesמצגת 8 ביוכימיה יסודות ואנזימים 2022Irina KniazevNo ratings yet
- כמה מים יש בהידראט- מתוקןDocument11 pagesכמה מים יש בהידראט- מתוקןElianna DemirjianNo ratings yet
- מבחן מועד אDocument8 pagesמבחן מועד אarield69No ratings yet
- סיכום מבוא לאקולוגיהDocument8 pagesסיכום מבוא לאקולוגיהapi-3831508100% (1)
- דוח מכין 1Document3 pagesדוח מכין 1piper27No ratings yet
- סכום העל בביוכימיהDocument64 pagesסכום העל בביוכימיהAlaa HajNo ratings yet
- נוסחאון ביוכימיה 2Document2 pagesנוסחאון ביוכימיה 2api-3806097No ratings yet
- מעבדה מספר 6 אופרון הלקטוזDocument2 pagesמעבדה מספר 6 אופרון הלקטוזitaykaNo ratings yet
- חמצון של ציקלוהקסנון לחומצה אדיפיתDocument1 pageחמצון של ציקלוהקסנון לחומצה אדיפיתKobi Angel SabanNo ratings yet
- 5 גליקוליזהDocument11 pages5 גליקוליזהירדן לויןNo ratings yet
- Pdfתרגיל בית 8 תגובות התמרה ואלימינציה מעודכןDocument2 pagesPdfתרגיל בית 8 תגובות התמרה ואלימינציה מעודכןdaniNo ratings yet
- שיעור 16 - קומפלקס פירובט דהידרוגנאז ומעגל קרבסDocument14 pagesשיעור 16 - קומפלקס פירובט דהידרוגנאז ומעגל קרבסtamar.razNo ratings yet
- דוח מעבדהDocument3 pagesדוח מעבדהariecurzon100% (1)
- סריגים מולקולריים, אטומריים, יוניים ומתכתיים ברמות הבנה שונותDocument10 pagesסריגים מולקולריים, אטומריים, יוניים ומתכתיים ברמות הבנה שונותaina1196No ratings yet
- דוח מכין קינטיקה מעבדה בכימיה כללית הפתוחהDocument4 pagesדוח מכין קינטיקה מעבדה בכימיה כללית הפתוחהFrostyWolfNo ratings yet
- דף נוסחאות ביוכימיה ב'Document3 pagesדף נוסחאות ביוכימיה ב'liaorNo ratings yet
- פיזיולוגיה שלי1Document25 pagesפיזיולוגיה שלי1Shahar Tam100% (3)
- 1-מבנה אלקטרוני וקישור+חומצות ובסיסיםDocument134 pages1-מבנה אלקטרוני וקישור+חומצות ובסיסיםMaya LigalemNo ratings yet
- תרגול 6Document19 pagesתרגול 6Shahar Tam100% (1)
- שחזור ביוכימיה מועד א' 2010Document6 pagesשחזור ביוכימיה מועד א' 2010Avi Hanna ManelaNo ratings yet
- Chemistrya 2021 9-10 TestDocument8 pagesChemistrya 2021 9-10 Testinbalsaa2008No ratings yet
- Exp. 2 Nucleophilic SusbstitutuionDocument11 pagesExp. 2 Nucleophilic Susbstitutuionzzjdzz100% (1)
- דוח מכין כימיה כללית מעבדה 6Document6 pagesדוח מכין כימיה כללית מעבדה 6zzjdzzNo ratings yet
- חלבוניםDocument11 pagesחלבוניםapi-25967322No ratings yet
- Help 10Document5 pagesHelp 10zzjdzzNo ratings yet
- ביולוגיה מולקולרית - שיעור 5 - 110308Document3 pagesביולוגיה מולקולרית - שיעור 5 - 110308api-3738386100% (1)
- דוח מסכם סטריאוכימיהDocument10 pagesדוח מסכם סטריאוכימיהmayah sabarNo ratings yet
- תשעא 17.02.2011 חורף א+פתרונותDocument9 pagesתשעא 17.02.2011 חורף א+פתרונותtalsw swissaaa100% (1)
- תרגיל בית 4 מנגנוני תגובה ותגובות אלקנים ואלקיניםDocument3 pagesתרגיל בית 4 מנגנוני תגובה ותגובות אלקנים ואלקיניםmayah sabarNo ratings yet
- סיכום קורס וירוסיםDocument29 pagesסיכום קורס וירוסיםShir DahanNo ratings yet
- 820Document318 pages820НиколайКарабаджак100% (3)
- Lab ManDocument39 pagesLab ManJenny KovaleveNo ratings yet
- ריאקציות של הבנזןDocument14 pagesריאקציות של הבנזןdrugon23No ratings yet
- כימיה כללית - 05.11.06Document4 pagesכימיה כללית - 05.11.06api-3738386No ratings yet
- שיעור 10 - מבוא לסוכריםDocument8 pagesשיעור 10 - מבוא לסוכריםtamar.razNo ratings yet
- מיקרוביולוגיה - שיעור 10 - 210508Document7 pagesמיקרוביולוגיה - שיעור 10 - 210508api-3738386No ratings yet
- הנדסה גנטית- מעודכןDocument11 pagesהנדסה גנטית- מעודכןMerav CohenNo ratings yet
- מעבדה קינטיקהDocument11 pagesמעבדה קינטיקהapi-3801666100% (1)
- ביוכימיה - שיעור מספר 3 - 07.03.07Document4 pagesביוכימיה - שיעור מספר 3 - 07.03.07api-37383860% (1)
- שיטות סמסטר ב סיכום1Document34 pagesשיטות סמסטר ב סיכום1Nitzan ArgamanNo ratings yet
- אימונולוגיה שיעור 2Document6 pagesאימונולוגיה שיעור 2Yael LifshitzNo ratings yet
- 3Document27 pages3אופיר גלNo ratings yet
- שיעור 7 19.3Document2 pagesשיעור 7 19.3tamar.razNo ratings yet
- בית חזהDocument73 pagesבית חזהtamar.razNo ratings yet
- שיעור 6 - המשך אנזימים וקינטיקת סטורציהDocument12 pagesשיעור 6 - המשך אנזימים וקינטיקת סטורציהtamar.razNo ratings yet
- ביוסינתזה של חומצות אמינו 2.4.20Document3 pagesביוסינתזה של חומצות אמינו 2.4.20tamar.razNo ratings yet
- מנגנוני טרנספורטDocument39 pagesמנגנוני טרנספורטtamar.razNo ratings yet
- - - סיכום אנטומיה - עמית מנור - בטן - מעודכן 270320Document17 pages- - סיכום אנטומיה - עמית מנור - בטן - מעודכן 270320tamar.raz100% (1)
- 4- אנזימיםDocument16 pages4- אנזימיםtamar.razNo ratings yet
- למידה עצמית עצבוב בית החזה, ניקוז לימפטיDocument11 pagesלמידה עצמית עצבוב בית החזה, ניקוז לימפטיtamar.razNo ratings yet
- שיעור 21 - המשך חימצון חומצות שומות, גופי קטון וביוסניתזה של חומצות שומןDocument10 pagesשיעור 21 - המשך חימצון חומצות שומות, גופי קטון וביוסניתזה של חומצות שומןtamar.razNo ratings yet
- סיכום יערה- ביולוגיה של התאDocument83 pagesסיכום יערה- ביולוגיה של התאtamar.razNo ratings yet
- שיעור 1- מבוא - התא- שיטות מחקרDocument2 pagesשיעור 1- מבוא - התא- שיטות מחקרtamar.razNo ratings yet
- ביולוגיה של התא- לוטם קוןDocument175 pagesביולוגיה של התא- לוטם קוןtamar.raz100% (1)
- ביוכימיה - עד ליפידיםDocument149 pagesביוכימיה - עד ליפידיםtamar.razNo ratings yet
- סיכומון ביוכימיה ללא קרישה 2025Document84 pagesסיכומון ביוכימיה ללא קרישה 2025tamar.raz100% (2)
- שיעור 5Document2 pagesשיעור 5tamar.razNo ratings yet
- שיעור 9 - ביואנרגיהDocument12 pagesשיעור 9 - ביואנרגיהtamar.razNo ratings yet
- 1 - חומצות אמינוDocument4 pages1 - חומצות אמינוtamar.razNo ratings yet
- שיעור 10 - מבוא לסוכריםDocument8 pagesשיעור 10 - מבוא לסוכריםtamar.razNo ratings yet
- 3 - מבנה החלבוןDocument15 pages3 - מבנה החלבוןtamar.razNo ratings yet
- ביופיזיקה - מאוחד (אופיר)Document75 pagesביופיזיקה - מאוחד (אופיר)tamar.razNo ratings yet
- גזים (מצגת 5)Document5 pagesגזים (מצגת 5)tamar.raz100% (1)
- בית חזה - הילה מאי - שחר דקלDocument79 pagesבית חזה - הילה מאי - שחר דקלtamar.razNo ratings yet
- שיעור 16 - קומפלקס פירובט דהידרוגנאז ומעגל קרבסDocument14 pagesשיעור 16 - קומפלקס פירובט דהידרוגנאז ומעגל קרבסtamar.razNo ratings yet
- שיעור 11- קליטת סוכרים במעי וטרנספורטרים לגלוקוזDocument6 pagesשיעור 11- קליטת סוכרים במעי וטרנספורטרים לגלוקוזtamar.razNo ratings yet
- כוחות בין מולקולריים (מצגת 9)Document7 pagesכוחות בין מולקולריים (מצגת 9)tamar.raz100% (1)
- מבנה האטום (מצגת 7)Document8 pagesמבנה האטום (מצגת 7)tamar.razNo ratings yet
- סיכום ביופיזיקה שיר ולבר וטל קולרDocument150 pagesסיכום ביופיזיקה שיר ולבר וטל קולרtamar.razNo ratings yet
- בית חזה - עמית יבורDocument75 pagesבית חזה - עמית יבורtamar.razNo ratings yet
- שיעור 2 אפידימולוגיה מדדי תחלואה ותמותה אמבר 4.11.19Document6 pagesשיעור 2 אפידימולוגיה מדדי תחלואה ותמותה אמבר 4.11.19tamar.razNo ratings yet