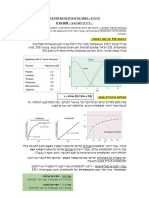
You might also like
- דו''ח 3Document10 pagesדו''ח 3Ksenia Khantaev0% (1)
- מעבדה במיקרוביולוגיה סניטרית 17.09.08Document112 pagesמעבדה במיקרוביולוגיה סניטרית 17.09.08hila220% (1)
- קובץ מושגים לקורס ביולוגיה מולקולרית וביטוי גניםDocument8 pagesקובץ מושגים לקורס ביולוגיה מולקולרית וביטוי גניםapi-3824802No ratings yet
- עזוב אותי באימש'ך-דוח 4Document21 pagesעזוב אותי באימש'ך-דוח 4api-3801666No ratings yet
- שאלות אמריקאיות בכימיה כלליתDocument4 pagesשאלות אמריקאיות בכימיה כלליתdrugon230% (1)
- דוח מעבדה 3 בדיקת גורמים המשפיעים על ריאקציה אנזימטית של האנזים פוספטאזטז חומצי (Acid phosphatase)Document8 pagesדוח מעבדה 3 בדיקת גורמים המשפיעים על ריאקציה אנזימטית של האנזים פוספטאזטז חומצי (Acid phosphatase)אנונימיNo ratings yet
- שאלות מבחן לדוגמא - ביוכימיה ומנוגנוני וויסותDocument3 pagesשאלות מבחן לדוגמא - ביוכימיה ומנוגנוני וויסותMariana SaadaNo ratings yet
- סיכום לקורס ביולוגיה מולקולרית וביטוי גנים תשסגDocument31 pagesסיכום לקורס ביולוגיה מולקולרית וביטוי גנים תשסגapi-3824802100% (2)
- NoName 04052020071546Document5 pagesNoName 04052020071546Bayan HamedNo ratings yet
- ביולוגיה מולקולרית פתרונותDocument19 pagesביולוגיה מולקולרית פתרונותYasmin NishriNo ratings yet
- נוסחאון ביוכימיה 2Document2 pagesנוסחאון ביוכימיה 2api-3806097No ratings yet
- דוח הגשה ביו-2Document7 pagesדוח הגשה ביו-2api-3801666No ratings yet
- שחזור ביוכימיה שלנוDocument13 pagesשחזור ביוכימיה שלנוapi-25967322100% (1)
- דוח 5Document14 pagesדוח 5api-3831508100% (4)
- גנטיקה - שיעור 13 - 021207Document5 pagesגנטיקה - שיעור 13 - 021207api-3738386No ratings yet
- מעבדה מספר 6 אופרון הלקטוזDocument2 pagesמעבדה מספר 6 אופרון הלקטוזitaykaNo ratings yet
- Do7 5Document6 pagesDo7 5khaledbek1No ratings yet
- - - סיכום אנטומיה - עמית מנור - בטן - מעודכן 270320Document17 pages- - סיכום אנטומיה - עמית מנור - בטן - מעודכן 270320tamar.raz100% (1)
- סיכומון ביוכימיה ללא קרישה 2025Document84 pagesסיכומון ביוכימיה ללא קרישה 2025tamar.raz100% (2)
- מנגנוני טרנספורטDocument39 pagesמנגנוני טרנספורטtamar.razNo ratings yet
- סיכום יערה- ביולוגיה של התאDocument83 pagesסיכום יערה- ביולוגיה של התאtamar.razNo ratings yet
- שיעור 16 - קומפלקס פירובט דהידרוגנאז ומעגל קרבסDocument14 pagesשיעור 16 - קומפלקס פירובט דהידרוגנאז ומעגל קרבסtamar.razNo ratings yet
- כוחות בין מולקולריים (מצגת 9)Document7 pagesכוחות בין מולקולריים (מצגת 9)tamar.raz100% (1)
- ביוכימיה4Document4 pagesביוכימיה4Zohar ShaharabaniNo ratings yet
- תרגול 4Document62 pagesתרגול 4Shahar TamNo ratings yet
- מעבדהDocument13 pagesמעבדהEmad AwadNo ratings yet
- ביוכימיה שירDocument236 pagesביוכימיה שירירדן לויןNo ratings yet
- וכימה כללית וקלינית-אנזימיםDocument23 pagesוכימה כללית וקלינית-אנזימיםapi-25967322100% (1)
- סיכום משופר ביוכימיהDocument34 pagesסיכום משופר ביוכימיהrichter1No ratings yet
- ביוכימיה5Document8 pagesביוכימיה5Zohar ShaharabaniNo ratings yet
- ביוכימיה של חלבונים סיכומיםDocument23 pagesביוכימיה של חלבונים סיכומיםannaerel100% (1)
- דף ביוכימיה-שירליDocument9 pagesדף ביוכימיה-שירליAlaa HajNo ratings yet
- מצגת 8 ביוכימיה יסודות ואנזימים 2022Document115 pagesמצגת 8 ביוכימיה יסודות ואנזימים 2022Irina KniazevNo ratings yet
- סיכום אנזימיםDocument3 pagesסיכום אנזימיםrachel112100% (1)
- שחזור ביוכימיה מועד א' 2010Document6 pagesשחזור ביוכימיה מועד א' 2010Avi Hanna ManelaNo ratings yet
- סיכומים בביוכימיהDocument3 pagesסיכומים בביוכימיהapi-3717496No ratings yet
- חלבוניםDocument11 pagesחלבוניםapi-25967322No ratings yet
- ביולוגיה מולקולרית 2018 PDFDocument95 pagesביולוגיה מולקולרית 2018 PDFYasmin AdarNo ratings yet
- דוח מסכם סטריאוכימיהDocument10 pagesדוח מסכם סטריאוכימיהmayah sabarNo ratings yet
- ביולוגיה מולקולרית - שיעור 5 - 110308Document3 pagesביולוגיה מולקולרית - שיעור 5 - 110308api-3738386100% (1)
- מיקרוביולוגיה סיכומונהDocument148 pagesמיקרוביולוגיה סיכומונהמירי צבריNo ratings yet
- מעבדה 6 ביוכימיהDocument8 pagesמעבדה 6 ביוכימיהStav LeviNo ratings yet
- דוח מעבדה בהנדסה גנטית - שיבוט וטרנספורמציהDocument10 pagesדוח מעבדה בהנדסה גנטית - שיבוט וטרנספורמציהAmir Erel100% (1)
- שיעור חזרה בגנטיקהDocument78 pagesשיעור חזרה בגנטיקהמשתמשת משתשמתNo ratings yet
- סכום העל בביוכימיהDocument64 pagesסכום העל בביוכימיהAlaa HajNo ratings yet
- מבחנים שנה ב אביב 2008- חוברת אגודהDocument490 pagesמבחנים שנה ב אביב 2008- חוברת אגודהapi-3740505No ratings yet
- גנטיקהDocument36 pagesגנטיקהnatigabi100% (1)
- שחזור בגנטיקה 2008Document3 pagesשחזור בגנטיקה 2008api-25967322No ratings yet
- ביוכימיה - עד ליפידיםDocument149 pagesביוכימיה - עד ליפידיםtamar.razNo ratings yet
- דף עזר שחרDocument15 pagesדף עזר שחרShahar TamNo ratings yet
- - - חוברת מעבדה-מיקרוביולוגיה-20Document33 pages- - חוברת מעבדה-מיקרוביולוגיה-20saja MuhammadNo ratings yet
- מעבדה אחרונה וסופיתDocument5 pagesמעבדה אחרונה וסופיתapi-3801666No ratings yet
- דוח מכין כימיה כללית מעבדה 6Document6 pagesדוח מכין כימיה כללית מעבדה 6zzjdzzNo ratings yet
- מיקרוביולוגיה PDFDocument18 pagesמיקרוביולוגיה PDFOrpaz MashkatoNo ratings yet
- ביוכימיה4מוכןDocument7 pagesביוכימיה4מוכןapi-3801666No ratings yet
- סיכום גניטיקהDocument29 pagesסיכום גניטיקהמשתמשת משתשמתNo ratings yet
- שאלות הכנה 3Document2 pagesשאלות הכנה 3api-3831508No ratings yet
- 820Document318 pages820НиколайКарабаджак100% (3)
- דוח סופיDocument32 pagesדוח סופיkarin_yanivNo ratings yet
- מיקרוביולוגיה2 סיכומונהDocument86 pagesמיקרוביולוגיה2 סיכומונהמירי צבריNo ratings yet
- ביוכימיה-דוח 1Document12 pagesביוכימיה-דוח 1api-38016660% (1)
- כימיה כללית - 30.11.06Document3 pagesכימיה כללית - 30.11.06api-3738386No ratings yet
- הקורס הנדסה גנטיתDocument44 pagesהקורס הנדסה גנטיתKeren LeviNo ratings yet
- דוח מעבדהDocument3 pagesדוח מעבדהariecurzon100% (1)
- Lab 2 MicrobiologyDocument15 pagesLab 2 MicrobiologyAnna MihaliovNo ratings yet
- שיטות סמסטר ב סיכום1Document34 pagesשיטות סמסטר ב סיכום1Nitzan ArgamanNo ratings yet
- שיעור 6 - המשך אנזימים וקינטיקת סטורציהDocument12 pagesשיעור 6 - המשך אנזימים וקינטיקת סטורציהtamar.razNo ratings yet
- בית חזהDocument73 pagesבית חזהtamar.razNo ratings yet
- שיעור 7 19.3Document2 pagesשיעור 7 19.3tamar.razNo ratings yet
- ביוסינתזה של חומצות אמינו 2.4.20Document3 pagesביוסינתזה של חומצות אמינו 2.4.20tamar.razNo ratings yet
- למידה עצמית עצבוב בית החזה, ניקוז לימפטיDocument11 pagesלמידה עצמית עצבוב בית החזה, ניקוז לימפטיtamar.razNo ratings yet
- שיעור 21 - המשך חימצון חומצות שומות, גופי קטון וביוסניתזה של חומצות שומןDocument10 pagesשיעור 21 - המשך חימצון חומצות שומות, גופי קטון וביוסניתזה של חומצות שומןtamar.razNo ratings yet
- ביוכימיה - עד ליפידיםDocument149 pagesביוכימיה - עד ליפידיםtamar.razNo ratings yet
- ביולוגיה של התא- לוטם קוןDocument175 pagesביולוגיה של התא- לוטם קוןtamar.raz100% (1)
- בית חזה - הילה מאי - שחר דקלDocument79 pagesבית חזה - הילה מאי - שחר דקלtamar.razNo ratings yet
- שיעור 5Document2 pagesשיעור 5tamar.razNo ratings yet
- שיעור 1- מבוא - התא- שיטות מחקרDocument2 pagesשיעור 1- מבוא - התא- שיטות מחקרtamar.razNo ratings yet
- 2 - שיטות ניתוח חלבוניםDocument10 pages2 - שיטות ניתוח חלבוניםtamar.razNo ratings yet
- שיעור 9 - ביואנרגיהDocument12 pagesשיעור 9 - ביואנרגיהtamar.razNo ratings yet
- 4- אנזימיםDocument16 pages4- אנזימיםtamar.razNo ratings yet
- שיעור 11- קליטת סוכרים במעי וטרנספורטרים לגלוקוזDocument6 pagesשיעור 11- קליטת סוכרים במעי וטרנספורטרים לגלוקוזtamar.razNo ratings yet
- 1 - חומצות אמינוDocument4 pages1 - חומצות אמינוtamar.razNo ratings yet
- 3 - מבנה החלבוןDocument15 pages3 - מבנה החלבוןtamar.razNo ratings yet
- בית חזה - עמית יבורDocument75 pagesבית חזה - עמית יבורtamar.razNo ratings yet
- שיעור 10 - מבוא לסוכריםDocument8 pagesשיעור 10 - מבוא לסוכריםtamar.razNo ratings yet
- סיכום ביופיזיקה שיר ולבר וטל קולרDocument150 pagesסיכום ביופיזיקה שיר ולבר וטל קולרtamar.razNo ratings yet
- גזים (מצגת 5)Document5 pagesגזים (מצגת 5)tamar.raz100% (1)
- מבנה האטום (מצגת 7)Document8 pagesמבנה האטום (מצגת 7)tamar.razNo ratings yet
- ביופיזיקה - מאוחד (אופיר)Document75 pagesביופיזיקה - מאוחד (אופיר)tamar.razNo ratings yet
- שיעור 2 אפידימולוגיה מדדי תחלואה ותמותה אמבר 4.11.19Document6 pagesשיעור 2 אפידימולוגיה מדדי תחלואה ותמותה אמבר 4.11.19tamar.razNo ratings yet