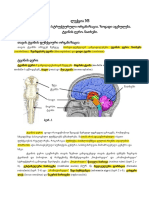
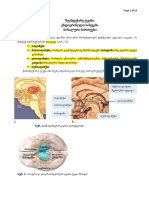
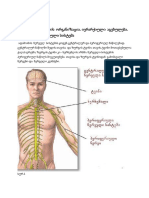
You might also like
- 158733Document9 pages158733Lizz WeasleyNo ratings yet
- 180690Document4 pages180690Nika AbashidzeNo ratings yet
- ნეირონი, როგორც მთავარი ნერვული უჯრედი. გლიაDocument9 pagesნეირონი, როგორც მთავარი ნერვული უჯრედი. გლიაNika AbashidzeNo ratings yet
- 3ნერვული უჯრედების წარმოშობა, ზრდა და გადარჩენაDocument6 pages3ნერვული უჯრედების წარმოშობა, ზრდა და გადარჩენაNika AbashidzeNo ratings yet
- პირველი ლექცია სკრიპტიDocument27 pagesპირველი ლექცია სკრიპტიTea GocadzeNo ratings yet
- Neiro Savalalo KonspektiDocument42 pagesNeiro Savalalo KonspektiLizi-tamari TsertsvadzeNo ratings yet
- 6 Jun 1 2docx PDF FreeDocument361 pages6 Jun 1 2docx PDF Freeთემო ვაგიევიNo ratings yet
- ფიზიოლოგია 2Document4 pagesფიზიოლოგია 244p52gtkbnNo ratings yet
- მე 6 თავიDocument23 pagesმე 6 თავიNinooo ONo ratings yet
- Orca Share Media1677930088774 7037748883065182885-1Document5 pagesOrca Share Media1677930088774 7037748883065182885-1Ketevani KhuroshviliNo ratings yet
- ნერვული სისტემის კონსპექტიDocument5 pagesნერვული სისტემის კონსპექტიDachi DavitashviliNo ratings yet
- ნერვული სისტემა -Document16 pagesნერვული სისტემა -mariamiNo ratings yet
- ნერვული სისტემის პათოლოგიაDocument8 pagesნერვული სისტემის პათოლოგიაSalome GvelesianiNo ratings yet
- Biochemistry ?Document65 pagesBiochemistry ?Anita PatieshviliNo ratings yet
- BiochemistryDocument65 pagesBiochemistryz27qj9n7f4No ratings yet
- ნატალია მხითარიანიDocument5 pagesნატალია მხითარიანიMariam LobjanidzeNo ratings yet
- ლექცია 2Document7 pagesლექცია 2Tika KurtanidzeNo ratings yet
- Fiziologia 1Document114 pagesFiziologia 1ანა მიქავაNo ratings yet
- 180688Document47 pages180688Nika AbashidzeNo ratings yet
- ბიოქიმია ნ2Document7 pagesბიოქიმია ნ2Saba BeruchashviliNo ratings yet
- Nucleus Ruber)Document9 pagesNucleus Ruber)Giorgi BarnabishviliNo ratings yet
- ფიზიო 3Document7 pagesფიზიო 3Ana UsharidzeNo ratings yet
- ნერვიDocument37 pagesნერვიsalome beridzeNo ratings yet
- ონტოგენეზი - განვითარების გენეტიკაDocument9 pagesონტოგენეზი - განვითარების გენეტიკაAnaNo ratings yet
- #1 ნერვული სისტემა, ნერვული ქსოვილიDocument14 pages#1 ნერვული სისტემა, ნერვული ქსოვილითეონა ბასილაძეNo ratings yet
- წიგნი - ნერვული სისტემაDocument102 pagesწიგნი - ნერვული სისტემაRatiNo ratings yet
- ადამიანის თავის ტვინი ყველაზე უფრო დახვეწილი და მაღალგანვითაDocument21 pagesადამიანის თავის ტვინი ყველაზე უფრო დახვეწილი და მაღალგანვითაana.turashvili056No ratings yet
- ქვიზი 1 ნევროლოგიაDocument6 pagesქვიზი 1 ნევროლოგიაLuka AbuladzeNo ratings yet
- 6Document12 pages6Giorgi Barnabishvili100% (1)
- მიტოზის ფაზებიDocument3 pagesმიტოზის ფაზებიMate Metreveli100% (1)
- კონსპექტი 2Document12 pagesკონსპექტი 2Gigi GiorgadzeNo ratings yet
- CNS - AnatomDocument42 pagesCNS - AnatomGiorgi LortkipanidzeNo ratings yet
- ბიოლოგიის ფინალურის ბილეთების პასუხები (შესწორებული) დ.გDocument30 pagesბიოლოგიის ფინალურის ბილეთების პასუხები (შესწორებული) დ.გKeti Kimadze100% (1)
- ბიოლოგიის კონსპექტი ბილეთებით.docx · version 1Document31 pagesბიოლოგიის კონსპექტი ბილეთებით.docx · version 1KaiNo ratings yet
- ბიოლოგია 2Document3 pagesბიოლოგია 2Vaska KoxashviliNo ratings yet
- Nervuli Sistema4Document67 pagesNervuli Sistema4nanukaNo ratings yet
- ნერვული სისტემა, ნერვული და გლიური უჯრედებიDocument43 pagesნერვული სისტემა, ნერვული და გლიური უჯრედებიGiorgi GafrindashviliNo ratings yet
- Nerve TissueDocument7 pagesNerve TissueTika TevdoradzeNo ratings yet
- 1. ნევროლოგიაDocument12 pages1. ნევროლოგიაAno DzamelashviliNo ratings yet
- გენომის ეპიგენეტიკური მოდელირება 1Document11 pagesგენომის ეპიგენეტიკური მოდელირება 1Tinatın TsıntsadzeNo ratings yet
- ადამიანის ემბრიოლოგიაDocument90 pagesადამიანის ემბრიოლოგიაAnita LeviNo ratings yet
- эмбриоDocument9 pagesэмбриоKristinaNo ratings yet
- эмбриоDocument9 pagesэмбриоKristinaNo ratings yet
- Download1 PDFDocument12 pagesDownload1 PDFNuca GvarakidzeNo ratings yet
- Download1 PDFDocument12 pagesDownload1 PDFNuca GvarakidzeNo ratings yet
- უჯრედის ონტოგენეზიDocument20 pagesუჯრედის ონტოგენეზიAnaNo ratings yet
- 7. უჯრედული კომუნიკაციებიDocument4 pages7. უჯრედული კომუნიკაციებიGiorgi RakviashviliNo ratings yet
- ნ ს -3Document5 pagesნ ს -3r2vxczxrp9No ratings yet
- ნერვული სისტემაDocument19 pagesნერვული სისტემაTika TevdoradzeNo ratings yet
- ნერვული რეგულაციის ძირითადი პრინციპებიDocument6 pagesნერვული რეგულაციის ძირითადი პრინციპებიNika AbashidzeNo ratings yet
- 1.1. ტვინის ევოლუციაDocument36 pages1.1. ტვინის ევოლუციაTsotne Arabuli100% (1)
- არაბირთვული გენომიDocument7 pagesარაბირთვული გენომიJaneNo ratings yet
- I - 4311 4304 4309 4312 - 4305 4312 4317 4314 4317 4306 4312 4304 - 1Document32 pagesI - 4311 4304 4309 4312 - 4305 4312 4317 4314 4317 4306 4312 4304 - 1Maria MaisuradzeNo ratings yet
- ნერვული სისტემის ელემენტებიDocument36 pagesნერვული სისტემის ელემენტებიCoralinaNo ratings yet
- ჰისტოDocument9 pagesჰისტო私の名前は ナナNo ratings yet
- 1. უჯრედი და გენომიDocument4 pages1. უჯრედი და გენომიMaria MaisuradzeNo ratings yet
- ქცევის ბიოლოგიური საფუძვლები. (სასწ. მასალა) თემა n4. თამარ ჯარიაშვილიDocument13 pagesქცევის ბიოლოგიური საფუძვლები. (სასწ. მასალა) თემა n4. თამარ ჯარიაშვილიNata KikvadzeNo ratings yet
- უჯრედის ბირთვიDocument13 pagesუჯრედის ბირთვიbacho onianiNo ratings yet
- 6Document8 pages6khatia khachidzeNo ratings yet
- 179934Document35 pages179934Nika AbashidzeNo ratings yet
- 180689Document9 pages180689Nika AbashidzeNo ratings yet
- American Journal of Health Promotion (3, 3 (1989) )Document68 pagesAmerican Journal of Health Promotion (3, 3 (1989) )Nika AbashidzeNo ratings yet
- 180688Document47 pages180688Nika AbashidzeNo ratings yet
- 85113Document70 pages85113Nika AbashidzeNo ratings yet
- 160222Document79 pages160222Nika AbashidzeNo ratings yet
- ბიოუსაფრთხოება და ბიოლოგიური დაცვა. ლ.11Document8 pagesბიოუსაფრთხოება და ბიოლოგიური დაცვა. ლ.11Nika Abashidze0% (1)
- საბჭოთა კავშირი - ლ - 6Document15 pagesსაბჭოთა კავშირი - ლ - 6Nika AbashidzeNo ratings yet
- ჟენევის 1925 წ კონფერენცია - ლ - 3Document12 pagesჟენევის 1925 წ კონფერენცია - ლ - 3Nika AbashidzeNo ratings yet
- საბჭოთა კავშირი - ლ - 6Document15 pagesსაბჭოთა კავშირი - ლ - 6Nika AbashidzeNo ratings yet
- ინგლისი - ლ - 7 2Document14 pagesინგლისი - ლ - 7 2Nika AbashidzeNo ratings yet
- AlkoholizmiDocument47 pagesAlkoholizmiNika AbashidzeNo ratings yet
- რა არის ბიოლოგიური იარაღი - ბი ადრეულ ხანებში ლ - 1Document15 pagesრა არის ბიოლოგიური იარაღი - ბი ადრეულ ხანებში ლ - 1Nika AbashidzeNo ratings yet
- იაპონიის პროგრამა 1931-1945 წლებში - ლ - 5Document13 pagesიაპონიის პროგრამა 1931-1945 წლებში - ლ - 5Nika AbashidzeNo ratings yet
- ბი - ლექცია2Document12 pagesბი - ლექცია2Nika AbashidzeNo ratings yet
- ბიოტერორიზმი და ბიოლოგიური თავდაცვა. ლ.10Document13 pagesბიოტერორიზმი და ბიოლოგიური თავდაცვა. ლ.10Nika AbashidzeNo ratings yet
- ამერიკის ბპ PDFDocument15 pagesამერიკის ბპ PDFNika AbashidzeNo ratings yet
- ბიოლოგიური ომის ისტორიის გაკვეთილები ლ.9Document15 pagesბიოლოგიური ომის ისტორიის გაკვეთილები ლ.9Nika AbashidzeNo ratings yet
- შეგრძნებაDocument16 pagesშეგრძნებაNika AbashidzeNo ratings yet
- ბიოტერორიზმი და ბიოლოგიური თავდაცვა. ლ.10Document13 pagesბიოტერორიზმი და ბიოლოგიური თავდაცვა. ლ.10Nika AbashidzeNo ratings yet
- ბი - ლექცია2Document12 pagesბი - ლექცია2Nika AbashidzeNo ratings yet
- რა არის ბიოლოგიური იარაღი - ბი ადრეულ ხანებში ლ - 1Document15 pagesრა არის ბიოლოგიური იარაღი - ბი ადრეულ ხანებში ლ - 1Nika AbashidzeNo ratings yet
- ბიოუსაფრთხოება და ბიოლოგიური დაცვა. ლ.11Document8 pagesბიოუსაფრთხოება და ბიოლოგიური დაცვა. ლ.11Nika Abashidze0% (1)
- ბიოტერორიზმი და ბიოლოგიური თავდაცვა. ლ.10Document13 pagesბიოტერორიზმი და ბიოლოგიური თავდაცვა. ლ.10Nika AbashidzeNo ratings yet
- ინგლისი - ლ - 7 2Document14 pagesინგლისი - ლ - 7 2Nika AbashidzeNo ratings yet
- ტკივილის ფენომენი და მისი ნეირონული საფუძვლებიDocument12 pagesტკივილის ფენომენი და მისი ნეირონული საფუძვლებიNika AbashidzeNo ratings yet
- ჟენევის 1925 წ კონფერენცია - ლ - 3Document12 pagesჟენევის 1925 წ კონფერენცია - ლ - 3Nika AbashidzeNo ratings yet
- იაპონიის პროგრამა 1931-1945 წლებში - ლ - 5Document13 pagesიაპონიის პროგრამა 1931-1945 წლებში - ლ - 5Nika AbashidzeNo ratings yet
- სიმღერა ნიბელუნგებზეDocument303 pagesსიმღერა ნიბელუნგებზეLuka BeqauriNo ratings yet