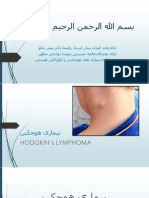
You might also like
- Cell Signaling and Post Translational ModificationDocument3 pagesCell Signaling and Post Translational ModificationGhazal HosseiniNo ratings yet
- 6702 2Document21 pages6702 2amal956No ratings yet
- Met Hapl AsiaDocument6 pagesMet Hapl AsiaNoor UllahNo ratings yet
- هورمون هاDocument125 pagesهورمون هاhamedhashimi100No ratings yet
- ویتامین های محلول در آبDocument65 pagesویتامین های محلول در آبbhjr98n7mnNo ratings yet
- ویتامین های محلول در آبDocument65 pagesویتامین های محلول در آبbhjr98n7mnNo ratings yet
- Capsules LectureDocument12 pagesCapsules Lectureshirzadnik7No ratings yet
- ایمونولوژیDocument10 pagesایمونولوژیdr.azimikitiNo ratings yet
- قارچ شناسی جلسه دومDocument6 pagesقارچ شناسی جلسه دومPegah SadeghiNo ratings yet
- Help SyndromeDocument13 pagesHelp SyndromenabizadadrsskNo ratings yet
- 4 6008339942055349427Document5 pages4 6008339942055349427sab4550aNo ratings yet
- Mammalian Polyamines - Non Canonical Base PairingDocument2 pagesMammalian Polyamines - Non Canonical Base PairingGhazal HosseiniNo ratings yet
- Medical Marijuana For Veterans Thesis StatementDocument30 pagesMedical Marijuana For Veterans Thesis StatementadibpeyrNo ratings yet
- HypersensitivityDocument15 pagesHypersensitivityarshamk keykhaeeNo ratings yet
- خون-جلسه 2Document10 pagesخون-جلسه 2AmethystNo ratings yet
- CossinDocument5 pagesCossinAli EslamiNo ratings yet
- آلکالوئیدها گیاه افدراDocument225 pagesآلکالوئیدها گیاه افدراAbbas NjjarkhatirNo ratings yet
- ?utf-8?B?2KjbjNmF2KfYsduM2YfYp9uMINin2LnYtdin2Kgg2Ygg2LHZiNin2YYu? ?utf-8?B?cHB0?Document33 pages?utf-8?B?2KjbjNmF2KfYsduM2YfYp9uMINin2LnYtdin2Kgg2Ygg2LHZiNin2YYu? ?utf-8?B?cHB0?sadaf babanezhadNo ratings yet
- CHEM Volume 4 Issue 13 Pages 68-74Document6 pagesCHEM Volume 4 Issue 13 Pages 68-74sheryNo ratings yet
- جلسه دوم پاتولوژی نظری کلیهDocument11 pagesجلسه دوم پاتولوژی نظری کلیهsalvatoredamon186444No ratings yet
- ترکیب اکسی کدون با آسپرین و استامینوفن و کدئین عالی ?????Document105 pagesترکیب اکسی کدون با آسپرین و استامینوفن و کدئین عالی ?????abasNo ratings yet
- تشوشات سیستم معافیتDocument7 pagesتشوشات سیستم معافیتMatin Hamid FaqiriNo ratings yet
- All1 1Document2 pagesAll1 1a.hosseiiinianNo ratings yet
- کودک بیمارDocument53 pagesکودک بیمارhyeju sonNo ratings yet
- فارمکولوژی ۳Document84 pagesفارمکولوژی ۳Awesome Videos100% (1)
- IJASR - Volume 9 - Issue 4 - Pages 400-412Document13 pagesIJASR - Volume 9 - Issue 4 - Pages 400-412omidNo ratings yet
- 4 5778433572515873852Document13 pages4 5778433572515873852dr.azimikitiNo ratings yet
- 7 هایپوکلسمیDocument15 pages7 هایپوکلسمیali mashhadiNo ratings yet
- Antidotes in Poisoning - PMCDocument11 pagesAntidotes in Poisoning - PMCAbbas NjjarkhatirNo ratings yet
- ChemotherapyDocument76 pagesChemotherapya.hosseiiinianNo ratings yet
- Evaluation of Cytotoxic Effects of Three Homeopathy Remedies (Sepia, Phosphorus and Pulsatilla) in The Breast Cancer Cell Line, MCF-7Document7 pagesEvaluation of Cytotoxic Effects of Three Homeopathy Remedies (Sepia, Phosphorus and Pulsatilla) in The Breast Cancer Cell Line, MCF-7Cat SkullNo ratings yet
- 200 پاینت مهم از پتالوژی عمومی رابنزDocument10 pages200 پاینت مهم از پتالوژی عمومی رابنزBashirNo ratings yet
- فصل نهمDocument30 pagesفصل نهمFattah GhulamiNo ratings yet
- فصل 39Document7 pagesفصل 39maliha arashNo ratings yet
- دارویی ورزشیDocument2 pagesدارویی ورزشیabbasiashkan97No ratings yet
- غده هیپوفیزDocument24 pagesغده هیپوفیزmohammad.hadi.mhvNo ratings yet
- PsoriasisDocument16 pagesPsoriasisSami ManiaNo ratings yet
- Mirhaji (Abstract, P)Document1 pageMirhaji (Abstract, P)majidNo ratings yet
- خون-جلسه 4Document7 pagesخون-جلسه 4AmethystNo ratings yet
- گزارش کار کاراموزیDocument24 pagesگزارش کار کاراموزیmhrm200020No ratings yet
- Womens Health Issues - حبیب زاده بیژنیDocument14 pagesWomens Health Issues - حبیب زاده بیژنیRazyNo ratings yet
- SYPHLISDocument20 pagesSYPHLISSakhawatullah SajadNo ratings yet
- 4 5814497652691176283Document55 pages4 5814497652691176283hiwaNo ratings yet
- Amir HoseiniDocument7 pagesAmir HoseiniAmir hoseiniNo ratings yet
- خون-جلسه 3Document10 pagesخون-جلسه 3AmethystNo ratings yet
- CNS2Document6 pagesCNS2ffNo ratings yet
- Multiple Sclerosis (M. S)Document38 pagesMultiple Sclerosis (M. S)ayazirezaNo ratings yet
- Diabetic Melitus PDFDocument39 pagesDiabetic Melitus PDFobaid.azizi.af1No ratings yet
- خون-جلسه 1Document12 pagesخون-جلسه 1AmethystNo ratings yet
- سندروم آنتی فسفولیپیدDocument8 pagesسندروم آنتی فسفولیپیدsabzavimaryam2001No ratings yet
- قارچ شناسیDocument6 pagesقارچ شناسیPegah SadeghiNo ratings yet
- Lecture 1 EndocrinologyDocument11 pagesLecture 1 EndocrinologySajidNo ratings yet
- Zist10 F2Document43 pagesZist10 F2Pouya VaeziNo ratings yet
- Therapeutic DiabeteDocument9 pagesTherapeutic DiabetearmanNo ratings yet
- پریزنتیشن درس هفتم تغذیDocument20 pagesپریزنتیشن درس هفتم تغذیAhmad MohammadiNo ratings yet
- سندرم X شکنندهDocument16 pagesسندرم X شکنندهAnonymous kVwp7DNo ratings yet
- LN 20180801111527 5455Document43 pagesLN 20180801111527 5455ATMNo ratings yet
- جزوه 4 درسنامه بیماری های اندوکراینDocument23 pagesجزوه 4 درسنامه بیماری های اندوکراینchnaper.hotmailNo ratings yet
- MineralDocument27 pagesMineralmuzhgan.sakhizada2008No ratings yet