You might also like
- 5.oksidatif FosforilasyonDocument46 pages5.oksidatif Fosforilasyonmusa karNo ratings yet
- Hücre Solunumu: Moleküler Biyoloji Ve Genetik BölümüDocument40 pagesHücre Solunumu: Moleküler Biyoloji Ve Genetik Bölümüq714514No ratings yet
- Oksi̇jenli̇ SolunumDocument6 pagesOksi̇jenli̇ SolunumMaksut TOSUNNo ratings yet
- Solun UmDocument66 pagesSolun Umkemalceba4No ratings yet
- Hafta13.Mikroorganizmaların Sınıflandırması Metabolizma SunumDocument34 pagesHafta13.Mikroorganizmaların Sınıflandırması Metabolizma SunumemirhanNo ratings yet
- 14 HaftaDocument19 pages14 HaftaHussem FelahNo ratings yet
- Hafta BiyoenejitikDocument14 pagesHafta Biyoenejitikşenol topuzNo ratings yet
- Hücresel SolunumDocument14 pagesHücresel SolunummychildhoodstoriesNo ratings yet
- 1.beslenme Biyokimyasına GirişDocument26 pages1.beslenme Biyokimyasına Giriş.No ratings yet
- FotosentezDocument40 pagesFotosentezEmrah ışıkNo ratings yet
- FotosentezDocument44 pagesFotosentezmusa karNo ratings yet
- Tca DöngüsüDocument99 pagesTca DöngüsüAytaç Gül50% (2)
- 10 Biyoenerjetik 1 DönüştürüldüDocument48 pages10 Biyoenerjetik 1 DönüştürüldüMustafa CingözNo ratings yet
- Hafta14 Katabolik-MetabolizmaDocument45 pagesHafta14 Katabolik-MetabolizmaemirhanNo ratings yet
- 13 HaftaDocument24 pages13 HaftaHussem FelahNo ratings yet
- 9 Hücresel SolunumDocument2 pages9 Hücresel Solunumsinem BoyalıNo ratings yet
- 3.karbohidrat Metabolizması, Sitrik Asit Çev.Document31 pages3.karbohidrat Metabolizması, Sitrik Asit Çev.musa karNo ratings yet
- 111 Biyokimya Ders 8Document66 pages111 Biyokimya Ders 8Yasin Çağrı KılıçerNo ratings yet
- CANLILARDA SOLUNUMDocument81 pagesCANLILARDA SOLUNUMBatu HanNo ratings yet
- Biyokimya ÖdevDocument5 pagesBiyokimya ÖdevlezzetNo ratings yet
- 5 EtzDocument14 pages5 EtzErhan KarabulutNo ratings yet
- Fotosentez-Oksi̇jenli̇ SolunumDocument15 pagesFotosentez-Oksi̇jenli̇ SolunumHIDIR ÇAĞRINo ratings yet
- Fotosentez Ders Notları1Document18 pagesFotosentez Ders Notları1merve şanserNo ratings yet
- 12 HaftaDocument23 pages12 HaftaHussem FelahNo ratings yet
- 2.9)sitrik asit döngüsüDocument75 pages2.9)sitrik asit döngüsünahzattamim360No ratings yet
- 7.karbonhi̇drat Metaboli̇zmasi (Kreps Döngüsü)Document38 pages7.karbonhi̇drat Metaboli̇zmasi (Kreps Döngüsü)Sarenur SoyturkNo ratings yet
- BiyoenerjetikDocument97 pagesBiyoenerjetikCeyhun YücelNo ratings yet
- 2008 2009+Biyokimya+II+Karbonhidrat+Met+2Document54 pages2008 2009+Biyokimya+II+Karbonhidrat+Met+2zuhalNo ratings yet
- Hücresel Solunum (Oksijenli Solunum)Document5 pagesHücresel Solunum (Oksijenli Solunum)nothing matterNo ratings yet
- Solunum Ve Fotosentez Karşilaştirma BüyükDocument4 pagesSolunum Ve Fotosentez Karşilaştirma Büyüktunagulsum58No ratings yet
- Karbon ReaksiyonlarıDocument49 pagesKarbon ReaksiyonlarıMoleküler BiyologNo ratings yet
- Biyokimya VDocument38 pagesBiyokimya VUğur ÇiğdemNo ratings yet
- 73) Oksijenli SolunumDocument10 pages73) Oksijenli SolunumA BNo ratings yet
- 1 BiyoenerjetiğinDocument40 pages1 Biyoenerjetiğinmusa karNo ratings yet
- BiyoanorganikDocument318 pagesBiyoanorganikspallocNo ratings yet
- Piruvat MetabolizmasıDocument26 pagesPiruvat Metabolizmasısean-paul38No ratings yet
- AntioksidanlarDocument56 pagesAntioksidanlarcerraheminNo ratings yet
- Fotosentez Solunum KemosentezDocument2 pagesFotosentez Solunum Kemosentezhilal -No ratings yet
- Ferment As YonDocument6 pagesFerment As YonMeliha GökmenNo ratings yet
- Biyokimya Ders 2 UKEYDocument35 pagesBiyokimya Ders 2 UKEYMousa AbazidNo ratings yet
- Polimerizasyon Ve Polimer Hakkında Genel BilgilerDocument95 pagesPolimerizasyon Ve Polimer Hakkında Genel Bilgilerqwert001No ratings yet
- 9 HaftaDocument21 pages9 HaftaHussem FelahNo ratings yet
- ENERJİ DÖNÜŞÜMLERİDocument25 pagesENERJİ DÖNÜŞÜMLERİBatu HanNo ratings yet
- 12 Sinif Ders Plani 2Document9 pages12 Sinif Ders Plani 2abarkindNo ratings yet
- Yağ Asidi Metabolizması PPDocument21 pagesYağ Asidi Metabolizması PPFatma AKATNo ratings yet
- Canlilari Geli̇şi̇m Süreci̇nde Enerji̇ Dönüşümü SonDocument54 pagesCanlilari Geli̇şi̇m Süreci̇nde Enerji̇ Dönüşümü Son9ayetokluNo ratings yet
- Ilaç Met 1Document174 pagesIlaç Met 1beyzakirtay5No ratings yet
- 06 BiyokimyaDocument19 pages06 Biyokimyaoncetus100% (1)
- BiyokimyaDocument11 pagesBiyokimyaistmNo ratings yet
- Karbonhidratlar (Şekerler)Document32 pagesKarbonhidratlar (Şekerler)arimbalimpetegimNo ratings yet
- Organi̇k Ki̇myaDocument12 pagesOrgani̇k Ki̇myaGamze SayarNo ratings yet
- Enerji SistemleriDocument60 pagesEnerji SistemleriSemaNo ratings yet
- 2018 FotosentezDocument52 pages2018 FotosentezBerker “Maniax88” TosunNo ratings yet
- Biyokimya Not - 2020 Tusdata Hizli TekrarDocument88 pagesBiyokimya Not - 2020 Tusdata Hizli TekrarBeyza Melike KarakaşNo ratings yet
- 1) Organi̇k Ki̇myaya Gi̇ri̇şDocument32 pages1) Organi̇k Ki̇myaya Gi̇ri̇şLARANo ratings yet
- Canlilarda Enerjy DonuyumleryDocument26 pagesCanlilarda Enerjy DonuyumlerymeldaNo ratings yet
- PROTEİNLER Son SonDocument26 pagesPROTEİNLER Son Sonezgigeyik02No ratings yet
- Sindirim BiyokimyasiDocument31 pagesSindirim Biyokimyasigülsüm önalNo ratings yet
- DIİmmunobiyolojiDocument8 pagesDIİmmunobiyolojigülsüm önalNo ratings yet
- Gözün Embriyolojik GelişimiDocument48 pagesGözün Embriyolojik Gelişimigülsüm önalNo ratings yet
- Kulakhisto Mayıs 2013Document61 pagesKulakhisto Mayıs 2013gülsüm önalNo ratings yet
- Kulağın Embriyolojik GelişimiDocument52 pagesKulağın Embriyolojik Gelişimigülsüm önalNo ratings yet
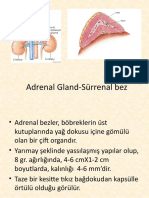
- Endokrin Sistem II HistolojisiDocument81 pagesEndokrin Sistem II Histolojisigülsüm önalNo ratings yet
- TusemDocument74 pagesTusemgülsüm önalNo ratings yet
- Tustime Deneme 62Document110 pagesTustime Deneme 62gülsüm önalNo ratings yet
- Göz HistolojisiDocument179 pagesGöz Histolojisigülsüm önalNo ratings yet
- Açıklamalı Cevaplar-1Document69 pagesAçıklamalı Cevaplar-1gülsüm önalNo ratings yet
- Tustime Deneme 61Document108 pagesTustime Deneme 61gülsüm önalNo ratings yet
- Tustime Deneme 63Document108 pagesTustime Deneme 63gülsüm önalNo ratings yet
- 1 Tustime (2017) PDFDocument72 pages1 Tustime (2017) PDFgülsüm önal100% (1)
- 5 Tusem (2017)Document56 pages5 Tusem (2017)gülsüm önalNo ratings yet
- 1-Baş KaslarıDocument3 pages1-Baş Kaslarıgülsüm önal100% (1)
- 2017 Tustime Deneme-59Document108 pages2017 Tustime Deneme-59gülsüm önalNo ratings yet
- 5 Tustime (2017)Document73 pages5 Tustime (2017)gülsüm önalNo ratings yet
- Norosirurjiozel6 3 3 PDFDocument7 pagesNorosirurjiozel6 3 3 PDFgülsüm önalNo ratings yet
- 2 Tustime (2017)Document74 pages2 Tustime (2017)gülsüm önalNo ratings yet
- Deneme Sınavı Soru DağılımıDocument64 pagesDeneme Sınavı Soru Dağılımıgülsüm önalNo ratings yet
- 3 Tusem (2017)Document64 pages3 Tusem (2017)gülsüm önalNo ratings yet
- 1 Tustime (2017) PDFDocument72 pages1 Tustime (2017) PDFgülsüm önal100% (1)
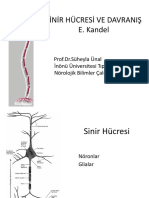
- Noroanatomi KandelDocument59 pagesNoroanatomi Kandelgülsüm önalNo ratings yet
- Noroanatomi KandelDocument59 pagesNoroanatomi Kandelgülsüm önalNo ratings yet
- Edebiyat Ve Tarih D - Nyas - Ndan Fizyoloji Ve T - P Alan - Ndaki Nobel - D - L - Ne - Eric Richard Kandel (#523823) - 643899Document10 pagesEdebiyat Ve Tarih D - Nyas - Ndan Fizyoloji Ve T - P Alan - Ndaki Nobel - D - L - Ne - Eric Richard Kandel (#523823) - 643899gülsüm önalNo ratings yet