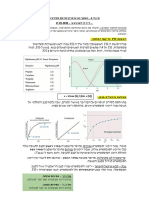
You might also like
- סיכומון ביוכימיה ללא קרישה 2025Document84 pagesסיכומון ביוכימיה ללא קרישה 2025tamar.raz100% (2)
- אנטומיה ופיסיולוגיה אDocument93 pagesאנטומיה ופיסיולוגיה אYael AbrahamiNo ratings yet
- פרמקולוגיה שיעור 3-4 פרמקודינמיקהDocument19 pagesפרמקולוגיה שיעור 3-4 פרמקודינמיקהOlya Pokotilo100% (1)
- ביולוגיה של התא-לימודי תואר ראשון של מדעי החיים- סופר סיכוםDocument41 pagesביולוגיה של התא-לימודי תואר ראשון של מדעי החיים- סופר סיכוםyuliakesler83% (6)
- הפקת GST ביוראקטוריםDocument21 pagesהפקת GST ביוראקטוריםOfir AmarNo ratings yet
- חומר טוב ביולוגיה של התאDocument50 pagesחומר טוב ביולוגיה של התאDaniel Shtriker67% (3)
- סיכום לקורס ביולוגיה מולקולרית וביטוי גנים תשסגDocument31 pagesסיכום לקורס ביולוגיה מולקולרית וביטוי גנים תשסגapi-3824802100% (2)
- מנגנוני טרנספורטDocument39 pagesמנגנוני טרנספורטtamar.razNo ratings yet
- - - סיכום אנטומיה - עמית מנור - בטן - מעודכן 270320Document17 pages- - סיכום אנטומיה - עמית מנור - בטן - מעודכן 270320tamar.raz100% (1)
- סיכומון ביוכימיה ללא קרישה 2025Document84 pagesסיכומון ביוכימיה ללא קרישה 2025tamar.raz100% (2)
- סיכום יערה- ביולוגיה של התאDocument83 pagesסיכום יערה- ביולוגיה של התאtamar.razNo ratings yet
- שיעור 16 - קומפלקס פירובט דהידרוגנאז ומעגל קרבסDocument14 pagesשיעור 16 - קומפלקס פירובט דהידרוגנאז ומעגל קרבסtamar.razNo ratings yet
- כוחות בין מולקולריים (מצגת 9)Document7 pagesכוחות בין מולקולריים (מצגת 9)tamar.raz100% (1)
- P1 Gpytmmb4Document36 pagesP1 Gpytmmb4לירן בוביNo ratings yet
- ביוכימיה - שיעור מספר 3 - 07.03.07Document4 pagesביוכימיה - שיעור מספר 3 - 07.03.07api-37383860% (1)
- סיכום פיזיולוגיה-אופירDocument273 pagesסיכום פיזיולוגיה-אופירrezq100% (1)
- שיעור 3- המשך שיטות הפרדת חלבונים ומבנה תלת מיימדיDocument16 pagesשיעור 3- המשך שיטות הפרדת חלבונים ומבנה תלת מיימדיtamar.razNo ratings yet
- חוברת אנטומיהDocument392 pagesחוברת אנטומיהGaia Shcherbina100% (1)
- אנדוקרינולוגיה - endocrinologyDocument46 pagesאנדוקרינולוגיה - endocrinologybar midamNo ratings yet
- פיזיולוגיה - הפיזיולוגיה של המערכת האינדוקרינית - 12912Document69 pagesפיזיולוגיה - הפיזיולוגיה של המערכת האינדוקרינית - 12912Alaa ShehadehNo ratings yet
- מעבדהDocument13 pagesמעבדהEmad AwadNo ratings yet
- וכימה כללית וקלינית-אנזימיםDocument23 pagesוכימה כללית וקלינית-אנזימיםapi-25967322100% (1)
- Wa0029 230202 232712Document66 pagesWa0029 230202 232712Lina AmouryNo ratings yet
- פיזיולוגיה של המערכות - 2014 - ליאורDocument269 pagesפיזיולוגיה של המערכות - 2014 - ליאורrezqNo ratings yet
- 1 - חומצות אמינוDocument4 pages1 - חומצות אמינוtamar.razNo ratings yet
- סיכום במדעי הצמחDocument59 pagesסיכום במדעי הצמחEinav Stern100% (1)
- 4- אנזימיםDocument16 pages4- אנזימיםtamar.razNo ratings yet
- שיעור 7Document6 pagesשיעור 7Nadia ReshiqNo ratings yet
- Copy of סיכום ביומולקולות 2016 - דניאל ויקטורDocument60 pagesCopy of סיכום ביומולקולות 2016 - דניאל ויקטורMeitar AtiasNo ratings yet
- 3Document27 pages3אופיר גלNo ratings yet
- פיזיולוגיה של מערכות - שיעור 8 - 010508Document11 pagesפיזיולוגיה של מערכות - שיעור 8 - 010508api-3738386100% (2)
- סיכום אנזימיםDocument3 pagesסיכום אנזימיםrachel112100% (1)
- ביוכימיה3Document4 pagesביוכימיה3Zohar ShaharabaniNo ratings yet
- אנדוקרינולוגיהDocument124 pagesאנדוקרינולוגיהBlich high School100% (3)
- דגם תאDocument10 pagesדגם תאHaim EzerNo ratings yet
- פרוטאזוםDocument1 pageפרוטאזוםapi-19979047No ratings yet
- 20Document3 pages20netaa6No ratings yet
- Bakanae ByoDocument21 pagesBakanae ByoBar AsulinNo ratings yet
- פרמקולוגיה מצגת מספר 1Document48 pagesפרמקולוגיה מצגת מספר 1Eldad Sarussi100% (1)
- הובלת תוך תאיתDocument7 pagesהובלת תוך תאיתapi-19979047No ratings yet
- Lesson15 30012008Document8 pagesLesson15 30012008api-25967322100% (2)
- Ac Files 472-1-1281 Introduction To Embryology and Histology 2010a H MFDocument45 pagesAc Files 472-1-1281 Introduction To Embryology and Histology 2010a H MFDror Dean BermanNo ratings yet
- הגשה 2Document6 pagesהגשה 2peterNo ratings yet
- ביוכימיה4Document4 pagesביוכימיה4Zohar ShaharabaniNo ratings yet
- תורת בלוטות מערכת ההפרשה הפנימיתDocument11 pagesתורת בלוטות מערכת ההפרשה הפנימיתrezqNo ratings yet
- תרגול 4Document62 pagesתרגול 4Shahar TamNo ratings yet
- פרופ' אמנון הופמן (גרסה 1) - סיכומיםDocument15 pagesפרופ' אמנון הופמן (גרסה 1) - סיכומיםSivan AndiNo ratings yet
- מצגת מעודכן 8Document22 pagesמצגת מעודכן 8AseelNo ratings yet
- סיכום מבנה ותפקוד של ביומולקולות - דר ראובן וינרDocument37 pagesסיכום מבנה ותפקוד של ביומולקולות - דר ראובן וינרMeitar AtiasNo ratings yet
- ביוכימיה א' דף שליף 3Document3 pagesביוכימיה א' דף שליף 3api-3831508100% (1)
- המחזור החודשי סיכום שיעורDocument4 pagesהמחזור החודשי סיכום שיעורapi-26159412No ratings yet
- סיכומים במדעיםDocument4 pagesסיכומים במדעיםItamar HenNo ratings yet
- הפרשה פנימית 1Document8 pagesהפרשה פנימית 1rezqNo ratings yet
- מאפייני החייםDocument3 pagesמאפייני החייםcarmitjulya228No ratings yet
- ביוכימיה5Document8 pagesביוכימיה5Zohar ShaharabaniNo ratings yet
- סיכום מאוחדDocument22 pagesסיכום מאוחדGal KahanaNo ratings yet
- ביוכימיה שירDocument236 pagesביוכימיה שירירדן לויןNo ratings yet
- מבנה התאDocument8 pagesמבנה התאSarah WeinbergerNo ratings yet
- אימונולוגיה שיעור 2Document6 pagesאימונולוגיה שיעור 2Yael LifshitzNo ratings yet
- סיכום זואולוגיהDocument83 pagesסיכום זואולוגיהSivanTakoNo ratings yet
- פרק 3 - התא הצמחיDocument5 pagesפרק 3 - התא הצמחיyuliakeslerNo ratings yet
- סכום הסטולוגיה2012Document36 pagesסכום הסטולוגיה2012Jasmin KolodroNo ratings yet
- סיכום משופר ביוכימיהDocument34 pagesסיכום משופר ביוכימיהrichter1No ratings yet
- ביוכימיה א חלק ב דרורDocument35 pagesביוכימיה א חלק ב דרורGal ZeevNo ratings yet
- גל וגלעדDocument27 pagesגל וגלעדapi-25967322No ratings yet
- שיעור 7 19.3Document2 pagesשיעור 7 19.3tamar.razNo ratings yet
- שיעור 6 - המשך אנזימים וקינטיקת סטורציהDocument12 pagesשיעור 6 - המשך אנזימים וקינטיקת סטורציהtamar.razNo ratings yet
- ביוסינתזה של חומצות אמינו 2.4.20Document3 pagesביוסינתזה של חומצות אמינו 2.4.20tamar.razNo ratings yet
- בית חזהDocument73 pagesבית חזהtamar.razNo ratings yet
- למידה עצמית עצבוב בית החזה, ניקוז לימפטיDocument11 pagesלמידה עצמית עצבוב בית החזה, ניקוז לימפטיtamar.razNo ratings yet
- שיעור 21 - המשך חימצון חומצות שומות, גופי קטון וביוסניתזה של חומצות שומןDocument10 pagesשיעור 21 - המשך חימצון חומצות שומות, גופי קטון וביוסניתזה של חומצות שומןtamar.razNo ratings yet
- ביוכימיה - עד ליפידיםDocument149 pagesביוכימיה - עד ליפידיםtamar.razNo ratings yet
- ביולוגיה של התא- לוטם קוןDocument175 pagesביולוגיה של התא- לוטם קוןtamar.raz100% (1)
- שיעור 1- מבוא - התא- שיטות מחקרDocument2 pagesשיעור 1- מבוא - התא- שיטות מחקרtamar.razNo ratings yet
- 2 - שיטות ניתוח חלבוניםDocument10 pages2 - שיטות ניתוח חלבוניםtamar.razNo ratings yet
- 3 - מבנה החלבוןDocument15 pages3 - מבנה החלבוןtamar.razNo ratings yet
- שיעור 9 - ביואנרגיהDocument12 pagesשיעור 9 - ביואנרגיהtamar.razNo ratings yet
- 4- אנזימיםDocument16 pages4- אנזימיםtamar.razNo ratings yet
- בית חזה - עמית יבורDocument75 pagesבית חזה - עמית יבורtamar.razNo ratings yet
- 1 - חומצות אמינוDocument4 pages1 - חומצות אמינוtamar.razNo ratings yet
- בית חזה - הילה מאי - שחר דקלDocument79 pagesבית חזה - הילה מאי - שחר דקלtamar.razNo ratings yet
- שיעור 11- קליטת סוכרים במעי וטרנספורטרים לגלוקוזDocument6 pagesשיעור 11- קליטת סוכרים במעי וטרנספורטרים לגלוקוזtamar.razNo ratings yet
- סיכום ביופיזיקה שיר ולבר וטל קולרDocument150 pagesסיכום ביופיזיקה שיר ולבר וטל קולרtamar.razNo ratings yet
- מבנה האטום (מצגת 7)Document8 pagesמבנה האטום (מצגת 7)tamar.razNo ratings yet
- שיעור 10 - מבוא לסוכריםDocument8 pagesשיעור 10 - מבוא לסוכריםtamar.razNo ratings yet
- גזים (מצגת 5)Document5 pagesגזים (מצגת 5)tamar.raz100% (1)
- ביופיזיקה - מאוחד (אופיר)Document75 pagesביופיזיקה - מאוחד (אופיר)tamar.razNo ratings yet
- שיעור 2 אפידימולוגיה מדדי תחלואה ותמותה אמבר 4.11.19Document6 pagesשיעור 2 אפידימולוגיה מדדי תחלואה ותמותה אמבר 4.11.19tamar.razNo ratings yet